Две французские косички. Варианты самостоятельного плетения.
Косичка-колосок — это актуальная повседневная прическа для девушек. Заплести 2 французские косички по бокам самой себе легче, чем кажется. Это аккуратная прическа, она может стать достойной альтернативой простому хвостику или пучку. Благодаря своей оригинальности она может быть повседневной или праздничной, такой вид укладки подойдет к любому наряду.
Правила плетения двух французских косичек поэтапноЧтобы с удобством заплести двойную французскую косу, понадобится небольшой набор вспомогательных материалов:
- Расческа-гребешок, оснащенная тонкой ручкой и частыми зубцами. Ею удобно разделять пряди на равные части.
- Простая расческа.
- Спрей для облегчения расчесывания.
- Резинки или ленты, заколки или иные аксессуары для прически.
- Фиксирующее средство (лак, мусс или гель).
Чтобы процесс плетения проходил без трудностей, есть некоторые правила для его облегчения:
- Перед тем как приступать к созданию колоска, требуется вымыть голову.

- На непослушные, склонные к запутыванию кудри, лучше нанести препарат, который упростит задачу.
- Пряди требуется тщательно расчесать перед началом.

Части для создания колоска делать одинаковыми, натягивать их равномерно.
Классический вариант колоскаЧтобы понять, как сделать две французские косички, требуется разобрать, что представляет собой одна. Это простая коса, но с вплетением отдельных прядок. Исполнить ее можно по следующему алгоритму:
- От прически отбирают группу прядей, которые еще используют для создания прически-мальвинки.
- Их делят на 3 равные части.
- Плетут простую косичку, в процессе прихватывая небольшие прядки с распущенных волос по бокам.
Когда все пряди будут задействованы в прическе, ее конец закрепляют резинкой или лентой.
Две французские косы — обратный колосокЭту прическу легко заплести, зная, как отрабатывается классический вариант. Эффект получившегося объема делает ее особенно изысканной. Процесс плетения 2 французских косичек «наоборот»:
Эффект получившегося объема делает ее особенно изысканной. Процесс плетения 2 французских косичек «наоборот»:
- Отбирают рабочую группу прядок, по бокам оставляют свободные волосы.
- В процессе плетения две внешние пряди кладут не на среднюю, а под нее (поэтому колосок называют обратным).
- С боковых частей равномерно вплетают небольшие прядки до полного задействования.
Так плетут один колосок. До того, как заплести две французские косы самой себе, прическу надо разделить:
- Колоски создают по очереди.
- С помощью мелкозубой расчески волосы аккуратно разбирают на две равные части.
- Чтобы они не мешали одна другой, требуется временно перехватить резинкой нерабочую половину.
- От второй отбирают три прядки, и заплетают классический колосок, захватывая свободные локоны. Когда он будет готов, завязывают на конце.
- Схему повторяют с другой стороны, не забывая о равномерном натяжении прядей для опрятности прически.
Плетение обеих косичек можно закончить в районе шеи для эффекта завершенности. Оставшиеся хвостики переплетают, объединяя две французских косы в цельную прическу.
Оставшиеся хвостики переплетают, объединяя две французских косы в цельную прическу.
Такой метод плетения 2 французских косичек отличается элегантностью и романтичностью. В каскад могут быть заплетены длинные или средние волосы. Сделать эту прическу можно по следующей схеме:
- Волосы расчесывают и слегка увлажняют. Можно применить мусс для фиксации.
- Разделяют рабочую область с обозначением пробора. Плетение начинается в височной части.
- Создают простой колосок.
- Делая пересечение прядей, нижняя опускается с захватом новой.
В завершение заплетают волосы по нужной длине.
Двойной пучок на короткие прядиОн хорошо ложится на волосы небольшой длины. Опрятно выглядит и поможет там, где нет возможности заплести вариант для средних или длинных прядей. Пошаговая инструкция плетения двух французских косичек:
- Причесывают и завивают при желании. Это сделает прическу пышнее.
- Волосы разделяют на три части. Средняя прядка для пучка, а внешние – для классических колосков.
- Заплетенные косички завязывают на конце и укладывают вокруг пучка.

Результат укрепляют заколками-невидимками или шпилькой.
Обруч из косичекЭто оригинальный метод создания прически на основе одной или двух французских кос. Хорошо дополняет даже строгий образ. Создание происходит по такой схеме:
- Голову причесывают.
- Выделяют часть локонов для плетения. Начинается оно от виска.
- Выделяют три пряди, из них создают косу, прихватывая свободные прядки. Их можно брать с лицевой стороны или с обоих, в зависимости от предпочитаемого метода.
- Косу заплетают до конца и прячут кончик с помощью невидимок.
Свободные концы можно завить.
Двойной «Рыбий хвост»Это простой метод плетения особого колоска. Отличается аккуратным, притягивающим взгляд видом. Плетение не занимает много времени, и выполняют его по следующему алгоритму. Как заплести две французские косички рыбкой:
Как заплести две французские косички рыбкой:
- Волосы расчесывают, разделяют надвое. Для этого применяют мелкозубую расческу с тонким хвостиком.
- Выделяют небольшую прядку с одной половины. Перекидывают ее на другую сторону.
- Это же проделывают с другой половиной, перекрещивают пряди.
Плетут такую косичку до конца и фиксируют резинкой.
Двойная объемная цепьЭто плетение укладывается на длинные и гладкие пряди аккуратнее, чем на другие типы. Основной аспект такой «цепочки» – простота исполнения. Выглядит готовая прическа, как ровная тонкая коса по центру, лежащая на других волосах. Пошаговая схема создания объемной цепочки для двух французских кос по бокам:
- Расчесывают.
- Обрабатывают утюжком.
- Зачесывают назад.
- Отбирают три тонких прядки — крайние от висков и одна в центре.
Плетут классическую французскую косу, прихватывая при этом еще и тонкие наружные пряди.
Французская коса, заплетенная по этому методу, выглядит празднично и элегантно. Она подходит для каждого типа волос. Придает эффект объема и густоты тонким волосам. Для облегчения плетения, голову требуется хорошо вымыть, тщательно расчесать. Можно применить спрей для причесывания и косметические препараты для укладки. Как самой себе заплести 2 французские косички:
- Волосы делят на 4 равные части. Первая прядь — слева, четвертая — справа.
- Третью прядь кладут поверх второй, контролируя центр. Он не должен сместиться.
- Четвертую накидывают на первую, зрительно она будет между третьей и первой.
- Вторую кладут под первую прядь. Сверху она закладывается третьей.
- Так проходит создание косы из четырех прядей. Алгоритму требуется внимательно следовать до завершения.
Есть упрощенный принцип плетения такой косы, который можно применить, если обычный вызывает затруднения. В нем 3 прядки с левой стороны плетут простым способом, последнюю постоянно кладут под крайнюю прядь справа.
В нем 3 прядки с левой стороны плетут простым способом, последнюю постоянно кладут под крайнюю прядь справа.
Двойные французские косы помогают создать образ для любого случая. Они нередко фигурируют на свадьбе в волосах невесты и у офисной леди ежедневно.
Такие прически придают женщине:
- элегантность;
- опрятность;
- красоту.
Вместе с колоском можно удачно использовать аксессуары:
- украшенные заколки;
- цветы;
- шпильки.
Носить такой вид укладки комфортно, косичка держит волосы в порядке, не мешает в быту.
Вариантов плетения существует большое количество. Это поможет каждый день выглядеть по-особенному. В плетении 2 французских косичек нет ничего трудного. Необходимо только понять принцип и попробовать.
Как научиться плести колосок?
Одним из признаков здоровых волос в древности считалась пышная и красивая девичья коса. Но сегодня очень мало девушек может похвастаться такой прической. Не смотря на это, девушка с длинными волосами, это бесспорно красиво.
Не смотря на это, девушка с длинными волосами, это бесспорно красиво.
Плести колоски или «французские косички» довольно легко. Многие девушки в школе делали их друг дружке. Если вы умели плести косички, то колоски у вас тоже будут получаться. Колосок – это, по сути, просто длинная коса, которую начинают плести с самой макушки. Сегодня есть много видов и техник плетения колосков, которые удивлят любых модниц. Но, не взирая на их сложность с первого взгляда, их легко сделать. Сегодня мы расскажем вам как это сделать.
Классический колосок
- Перед началом смочите водой волосы, совсем не много, иначе вы испортите колосок.
- Соберите волосы с боков, макушки, а также с передней части головы.
- Разделите волосы на 3 части. Начинать плести нужно как можно ближе к голове.
- Сначала вы должны сплести основную косу.
- После основной косы, добавляйте к пряди боковую прядь.
- Сплетите правую прядь (самую толстую) с средней прядью. Потом повторите с левой стороной. Продолжать следует до тех пор, пока не останется 10 см. После чего закрепите колосок ленточкой или резинкой. Так же, можно вплетать ленточку в волосы.
 Продолжать следует до тех пор, пока не останется 10 см. После чего закрепите колосок ленточкой или резинкой. Так же, можно вплетать ленточку в волосы.
Продолжать следует до тех пор, пока не останется 10 см. После чего закрепите колосок ленточкой или резинкой. Так же, можно вплетать ленточку в волосы.Другие варианты
- Вы можете экспериментировать, чтобы придать колоску разнообразия. У вас должны быть длинные волосы, а так же вы должны научиться плести обыкновенный колосок.
- Проще всего экспериментировать с 2-мя колосками. Вы можете украшать их, скреплять. Они плетутся как обычные, вам стоит просто довериться внутреннему чувству композиции.
- Так же можно соединить 2 колоска. Чтобы такое сделать, вам следует просто сплести два колоска по бокам головы, а потом соединить их в одну косу, либо в колосок.
- После того, как вы научитесь плести основные колоски, пробуйте придумывать собственные прически. Вы сможете заплетать их своим друзьям, родным.
Делаем из колосков рисунки
В зависимости от того, как именно на голове вы разместите колоски, вы можете изменять всю композицию прически и создавать свои рисунки.
- Можно заплести 3 колоска, лишь бы у вас хватило волос.
- Заплетите 6 очень маленьких колосков. Соберите их в некоторое подобие розочек. У вас получиться 3 розочки.
- Плести колоски можно и от середины головы. У вас получиться не обычная прическа. Оставшиеся волосы украсьте лентой.
Ищите, выдумывайте и творите. Все зависит от рамок вашей фантазии. Не имеет значения, какой рисунок из колосков вы сделаете. Главное, чтобы он смотрелся стильно и аккуратно.
Видео уроки
Плетём колосок
Вопрос о том, как плести колосок, возникает у большинства современных женщин, потому как данная прическа надежна, красива и затрачивает минимум времени.
Колосок был популярен еще 20 лет назад, однако затем его стали заменять более «свободными» прическами. Возможно это парадокс, но любая мода постепенно возвращается, преображаясь, благодаря новым тенденциям и веяниям.
Ранее колосок подразумевал аккуратность и идеальную гладкость, считаясь изящной, женственной прической, которая должна была соответствовать деловому строгому стилю.Сегодня же, напротив, при плетении колоска приветствуется легкая небрежность, и данная прическа великолепно вписывается в различные стили.
От обычной косы колосок отличается лишь более сложной техникой плетения. Но стоит немного потренироваться, как подобная коса сможет без усилий украшать женскую голову. Эта прическа прекрасно смотрится как на толстых, так и на тонких волосах. Разница в том, что при плетении колоска на тонких волосах лучше предварительно сделать небольшой начес – тогда прическа будет выглядеть объемно.
Плетение всех разновидностей колосков отталкивается от двух основных классических техник – «французской косы» и «рыбьего хвоста», которые мы и рассмотрим.
- Расчешите волосы. При необходимости сделайте легкий начес.
- Возьмите прядь волос со лба и разделите на три равные части.
- Начните заплетать обычную косу.
- Из волос, находящихся по бокам нужно поочередно — то справа, то слева вплетать дополнительные пряди.
- Чем вы чаще захватываете пряди с боков, тем колосок получается оригинальнее.
- Как правило, толщина дополнительных прядей определяется на глаз.
- Продолжайте плести колосок, пока дополнительные пряди не будут полностью вплетены в прическу.
- Оставшийся хвост можно закрепить резинкой или заплести обычную косу.
На сегодняшний день классический колосок претерпел массу трансформаций. Поэтому вы можете заплести не одну косу, а два колоска по бокам (фото 7) или, к примеру, несколько колосков, переходящих в более мелкие французские косички. Стоит иметь в виду, что современная мода приветствует колоски, заплетенные свободно. Это отчасти напоминает прическу, которую носят уже не первый день. При этом вовсе не обязательно вплетать все волосы. Для изюминки можно выпустить пару локонов, которые прекрасно обрамят лицо. Колосок, заплетенный по кругу вблизи ото лба, отлично заменит ободок, оставшиеся же волосы будут находиться в свободном падении. (фото 9) В какой именно части головы плести французскую косу выбирать только вам. Достаточно необычно выглядит колосок, заплетенный от основания шеи по направлению вверх. (фото 8) Оставшиеся пряди можно накрутить и создать роскошный пучок на макушке.
Стоит иметь в виду, что современная мода приветствует колоски, заплетенные свободно. Это отчасти напоминает прическу, которую носят уже не первый день. При этом вовсе не обязательно вплетать все волосы. Для изюминки можно выпустить пару локонов, которые прекрасно обрамят лицо. Колосок, заплетенный по кругу вблизи ото лба, отлично заменит ободок, оставшиеся же волосы будут находиться в свободном падении. (фото 9) В какой именно части головы плести французскую косу выбирать только вам. Достаточно необычно выглядит колосок, заплетенный от основания шеи по направлению вверх. (фото 8) Оставшиеся пряди можно накрутить и создать роскошный пучок на макушке.
- Зачешите все волосы назад. Если вы имеете челку, аккуратно отделите ее от остальных волос.
- Отделите пару тонких прядей волос, расположенных симметрично в области правого и левого виска.
- Перекрестите локоны
- Придерживая рукой получившееся скрещение, захватите следующую прядь с одного края и переместите ее на противоположную сторону. Тоже самое проделайте с другой стороны.
- Аналогичным образом продолжайте плетение до конца.
- Оставшиеся волосы закрепите резинкой или декоративной заколочкой.
 Тоже самое проделайте с другой стороны.
Тоже самое проделайте с другой стороны.Вам наверняка интересно узнать, как плести колосок, используя другие техники плетения? Тогда смотрите и изучайте. Безусловно, чтобы создавать такие прически потребуются месяца тренировок, но какой результат… Главное знать, как плести колосок посредством базовых техник, а дальше просто экспериментируйте и включайте больше фантазии.
Колоски на средние волосы: варианты плетения
Сегодня многие девушки любят косы. Они замечательно смотрятся, так как волосам придается красивая форма. Конечно, плетение кос – не совсем легкое занятие, и оно требует больших стараний. Чтобы получились шикарные и стильные прически на средние волосы, необходимо долго учиться и работать над этим. Но тем не менее время не будет потрачено зря, и в любой момент красивая прическа для выхода в свет будет готова. При этом не нужно будет посещать салоны и тратить деньги, ведь красоту можно сделать самостоятельно.
При этом не нужно будет посещать салоны и тратить деньги, ведь красоту можно сделать самостоятельно.
Стоит отметить, что косички всегда привлекают внимание людей, так как никогда не выходят из моды. Рассмотрим далее, как заплести колосок на средние волосы.
Подготовка
Чтобы прическа сохраняла свою свежесть на целый день, нужно очень старательно готовиться к ее выполнению. Самое главное – сразу помыть голову и смягчить волосы, используя бальзам. После этого нужно воспользоваться феном, но важно, чтобы поток воздуха из фена был холодным. Лучше всего сделать так, чтобы после мытья и применения фена волосы были немного влажные. Таким образом, будет удобнее плести косу.
Для создания причесок нужны будут следующие инструменты:
- резинки – тонкие и прочные, для фиксации причесок;
- невидимые заколки;
- различные украшения для разнообразия плетения кос;
- принадлежности для фиксации — лак, гель, мусс для придания дополнительного объема волосам.

Прическа «Корона»
С помощью правильно выполненного колоска на средние волосы можно создать необыкновенно красивый и эффектный образ.
Последовательность работы:
- Прежде всего, нужно хорошо расчесать волосы по всей их длине, чтобы отсутствовали запутанные пряди.
- Часть волос необходимо отделить. В первую очередь их требуется собрать от макушки до середины головы. Верхний слой завязывается резинкой, и хорошо, если она будет слегка заметной.
- Ниже середины головы волосы должны оставаться шириной в пять сантиметров.
- Верхний слой может быть подобен хвосту лошади и должен спадать вниз.
- Теперь можно начинать плести французскую косичку. Прядь нужно брать с левой стороны из нижнего слоя волос. И при укладке верхней части нужно забрать часть слоя из хвоста.
- Следует продолжать плетение, пока не будет достигнуто основание колоска.
- Прядь, которая осталась, заплетается в обычную косу, а конец волос связывается тонкой резинкой.
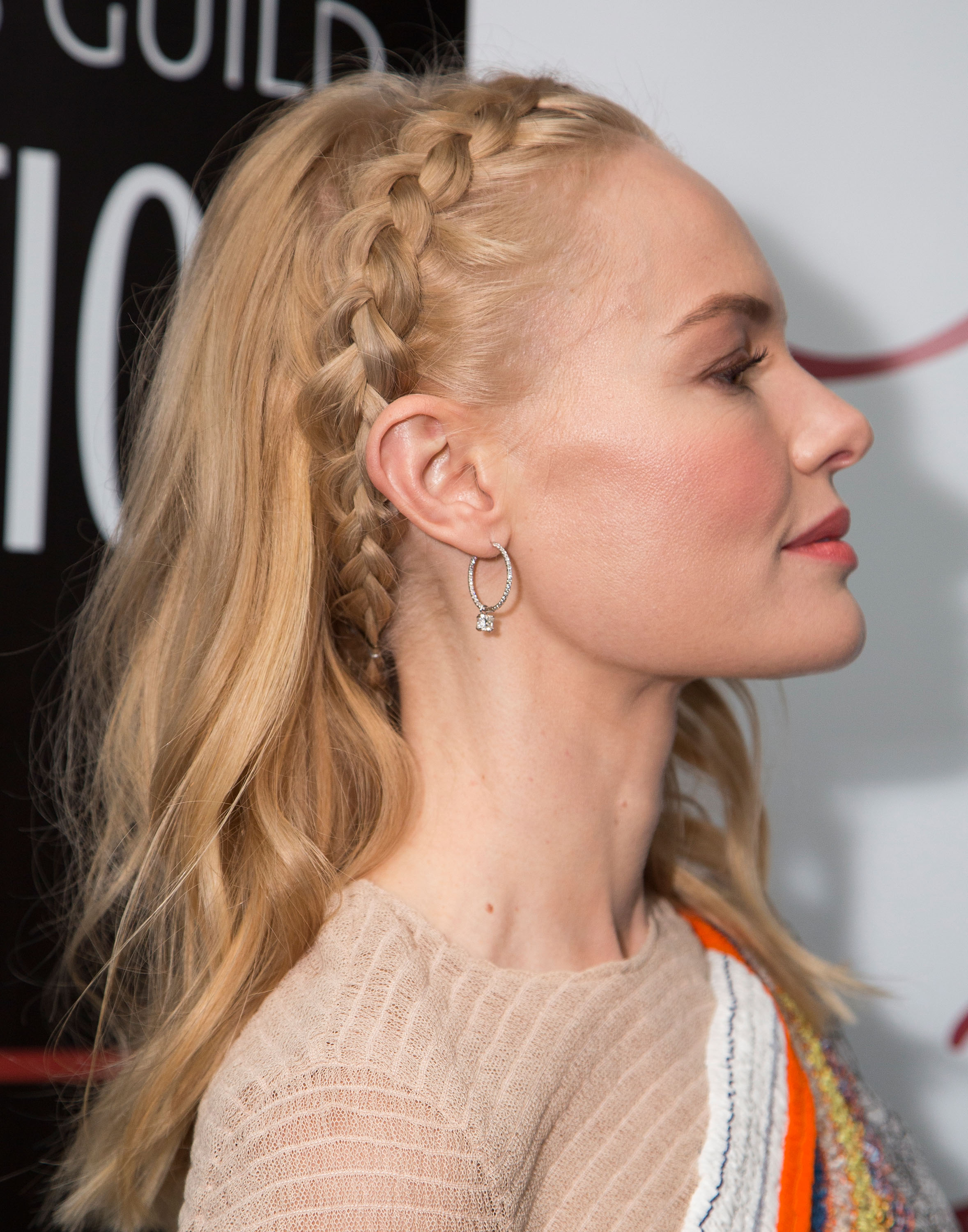
Для того чтобы скрыть косу, нужно тщательно завести ее вовнутрь короны, которая получилась. Чтобы она нормально держалась, ее также можно закрепить невидимками. Если все сделано правильно, то будет такое ощущение, что у колоска нет начала и конца. Также можно украсить этот колосок различными украшениями для большего эффекта.
Два колоска на средние волосы
Эта прическа отлично подходит не только взрослым девушкам, но и маленьким девочкам. Плетут два колоска так же, как и один. Изначально делают пробор, разделяя волосы на две части. Одну из них закрепляют заколкой или резинкой, чтобы она не мешала. Затем приступают непосредственно к самому плетению. Разделяют волосы на три одинаковые пряди. Поочередно кладут правую и левую прядь на центральную, увеличивая каждый раз ее объем за счет волос, оставшихся по бокам.
Прическа «Французская коса»
Такая прическа колосок представляет собой роскошный способ укладки средних волос. Она актуальна на все случаи жизни. Например, на свадьбу либо на романтический вечер такая прическа придется очень кстати.
Например, на свадьбу либо на романтический вечер такая прическа придется очень кстати.
Французская коса (колосок на средние волосы) имеет свою изюминку. В этом случае во время плетения могут быть использованы не все волосы, а только отдельные пряди маленького размера. Вариантов французских кос достаточно много, и любой можно создать как для повседневной жизни, так и на разные праздники. У прически много преимуществ. Самыми главными являются простота и своеобразие. Но все же нужно будет хорошо изучить технику плетения.
Последовательность плетения прически колосок:
- волосы надо расчесать и при этом использовать мусс для увеличения объема;
- вбок от лобной части необходимо отделить часть волос;
- пучок, который получился, надо связать маленькой резинкой;
- после этого можно потихоньку плести обычный колосок, который идет сбоку по росту волос;
- стоит учесть, что пряди волос по бокам постоянно должны быть уложены над серединной прядью в колоске;
- следует заплетать колосок до тех пор, пока боковые пряди не закончатся.

Для того чтобы волосы были объемными и прическа была очень красивая, через каждые три прохода необходимо взять прядь, которая заплетена, и потянуть ее в сторону. Тогда она будет похожа на кружево. Волосы, которые остались, желательно заплести в косу и подколоть особым украшением.
Прическа «Водопад»
Такую прическу колосок на средние волосы создать совсем просто. При этом она может создать особенный образ.
Последовательность работы:
- Начать плетение следует с лобной части. Прядь маленького размера отделяется и делится на 3 одинаковые доли.
- Из самой пряди нужно заплетать обычную косу, при этом сделав один проход.
- Во втором проходе можно добавить волосы сверху, опять же небольшую прядь, и продолжать плести.
- Как только две пряди сплетены между собой, нужно переплести большую прядь со средней.
- После всего этого из колоска убирается часть волос для того, чтобы они могли легко наклониться вниз.
- Сверху добавляются локоны, а после переплетения двух прядей их нужно убрать, при этом склоняя вниз.

После того как противоположная сторона достигнута, односторонний колосок заплетать больше не нужно, а просто доплести обычную косу до конца. Также следует завязать кончик резинкой.
Греческий стиль
Очень аккуратная и привлекательная прическа колосок на средние волосы. Она может произвести впечатление на любом мероприятии.
- Прежде всего, надо расчесать волосы и распределить боковой пробор с одной стороны.
- Чтобы пряди не мешали плетению, нужно заколоть часть кудрей набок.
- С левой стороны от пробора надо выделить маленькой пучок волос и разделить на три одинаковые части.
- Плетение необходимо начать с нижней части затылка и при этом вплетать пряди с правой стороны.
- Теперь нужно закрепить плетение заколкой и сделать такую же процедуру на другой стороне головы.
После всего надо закрепить две косы внизу затылка и убрать концы вовнутрь. Когда получился результат, можно обработать лаком волосы для придания дополнительного эффекта.
Прическа «Бохо стайл»
Много девушек предпочитают именно такой стиль прически, так как она вписывается в естественный образ и смотрится просто замечательно.
С помощью такого вида колоска на средние волосы можно разнообразить свой стиль. Особенно подходит бохо стайл тем, у кого прическа с косой челкой.
Последовательность плетения:
- Начать следует, прежде всего, с виска.
- Движение должно быть вдоль лба, по росту волос от одного до другого виска.
- Ободок, который получился около головы, нужно закрепить резинкой и заколками вверху. Желательно использовать незаметные заколки.
Прическа «Жгут»
Такая прическа используется достаточно часто, поскольку очень популярна среди девушек. Плести ее совершенно не сложно. Подойдет она больше всего как раз к волосам средней длины. Нужны будут только две пряди и немного терпения.
Плетение:
- Нужно расчесаться и собрать волосы в хвостик. При этом важно закрепить их резинкой.
- Разделить волосы на две равные части и скрутить каждую в жгутик.
- Потом нужно сплести эти жгуты между собой. В таком случае прическа будет подобна спирали.
- В конце завязать резинкой и использовать лак для волос.

Прическа «Дракон»
Создание такой прически не займет много времени, и к тому же, чтобы ее заплести, не стоит даже ходить в дорогие салоны. Она представляет собой обычный объемный колосок на средние волосы, но смотрится при этом очень интересно.
- плетение нужно начать постепенно, с центра макушки, двигаясь к шее;
- по бокам нужно собрать дополнительные пряди и завести их под низ предшествующих;
- коса, которая получилась, закрепляется резинкой либо же другими красивыми принадлежностями.
Четыре пряди
Иногда такое плетение кажется слишком сложным, но если проявить терпение, то все получится замечательно без больших усилий:
- первое, что нужно сделать – расчесать волосы и разделить на четыре части;
- завести первую часть волос за вторую, а третью завести за первую, сначала это кажется немного сложно, но это только в начале;
- четвертую прядь заложить под первую, а вторую – сверху на третью, также четвертую необходимо разместить поверх второй;
- продолжать проделывать такое плетение до самого окончания длины волос.

Сегодня существует огромное множество способов плетения самых разнообразных колосков на средние волосы. Научиться их плести совсем несложно. Варианты можно подбирать самые разные и украсить прическу можно интересными принадлежностями. У каждого свой вкус, и каждая девушка должна себе подбирать, то, что ей непременно по душе.
Научиться плести самой себе колоски на средние волосы несложно. В основном плетение кос используют для мероприятий или для проведения вечера в приятной компании, а в повседневной жизни девушки привыкли завязывать пряди в пучок либо вовсе распускать. Не стоит забывать, что ухоженные волосы смотрятся очень красиво и красят абсолютно всех девушек.
Как заплести колосок самой себе
Как заплести колосок самой себе
В наступающем сезоне самыми модными стали длинные волосы. Их носят распущенными, украсив заколками, обручами, диадемами, заплетают в косы и завязывают «конские» хвосты. В коллекциях модных дизайнеров представлены всевозможные плетения кос. Это и французская коса наоборот, и греческая коса, и плетение в виде корзиночек, и, конечно же, всеми любимая коса колосок. Колосок – это рыбий хвост Плетение колосок по-другому называется французской косой или рыбьим хвостом. Плетется он и на длинных волосах, и на волосах покороче, что очень важно для тех, кому хочется носить косы, но длина волос не позволяет это сделать. Колосок подходит для любого типа лица и любой структуры волос. Существует два вида плетения: колосок сбоку и рыбий хвост по центру головы. Как плести колосок самой себе — пошаговая инструкция Колосок по центру головы или рыбий хвост Вам потребуется расческа, резинка для волос и украшение по вкусу. Расчешите волосы назад, ото лба по направлению к затылку. Справа и слева от висков отделите небольшие тонкие пряди. Положите левую прядь под правую за головой. Держите правой рукой эти пряди, а левой возьмите слева еще одну прядку. Новую прядь пропустите под две пряди в правой руке. Теперь все пряди собрались в левой руке. Правой рукой отделяем прядку с правой стороны.
Это и французская коса наоборот, и греческая коса, и плетение в виде корзиночек, и, конечно же, всеми любимая коса колосок. Колосок – это рыбий хвост Плетение колосок по-другому называется французской косой или рыбьим хвостом. Плетется он и на длинных волосах, и на волосах покороче, что очень важно для тех, кому хочется носить косы, но длина волос не позволяет это сделать. Колосок подходит для любого типа лица и любой структуры волос. Существует два вида плетения: колосок сбоку и рыбий хвост по центру головы. Как плести колосок самой себе — пошаговая инструкция Колосок по центру головы или рыбий хвост Вам потребуется расческа, резинка для волос и украшение по вкусу. Расчешите волосы назад, ото лба по направлению к затылку. Справа и слева от висков отделите небольшие тонкие пряди. Положите левую прядь под правую за головой. Держите правой рукой эти пряди, а левой возьмите слева еще одну прядку. Новую прядь пропустите под две пряди в правой руке. Теперь все пряди собрались в левой руке. Правой рукой отделяем прядку с правой стороны. Переплетаем новую прядь с той, которая находится в левой руке. Повторяем отделение прядок поочередно с каждой стороны и переплетение их с основными прядями. Косу можно плести и до шеи, и до конца волос. На конце косы завяжите резинку. Украсьте прическу на свой вкус. Колосок сбоку
Переплетаем новую прядь с той, которая находится в левой руке. Повторяем отделение прядок поочередно с каждой стороны и переплетение их с основными прядями. Косу можно плести и до шеи, и до конца волос. На конце косы завяжите резинку. Украсьте прическу на свой вкус. Колосок сбоку
Боковые жгутики-косы выглядят очень оригинально. Плетутся они по принципу плетения французской косы, только каждая прядь, которую надо добавлять сбоку, перекручивается в виде жгута. Косы такого вида плетутся в сочетании с прямым пробором, который проходит по центру головы. Получается красивая прическа с двумя косами по бокам. Концы кос можно уложить в интересную «корзиночку» или оставить распущенными. Украшением такого колоска могут стать ленты, банты и заколки. Тем, кто заинтересовался плетением двух колосков, предлагается пошаговая инструкция, как заплести колосок самой себе с двух сторон головы. Вам потребуются: расческа, несколько простых шпилек, ленточка и резинка. Точно по центральной части головы делаем ровный пробор. Расчесываем волосы на две стороны. На каждой стороне должно быть одинаковое количество волос. Работаем с левой стороны. У виска от пробора отделяем тонкую прядь волос. Перекручиваем ее в сторону пробора. Держим жгут правой рукой. Левой рукой отделяем другую прядь. Она должна быть точно под первой. Делаем жгут из новой пряди. Переплетаем два жгута между собой, первый под вторым. Перекладываем оба жгута в правую руку и смешиваем в одну прядь. Повторяем пункты 4-8. Закрепляем концы волос под косой с помощью шпилек. Переходим к правой половине волос и повторяем все с начала. Под подогнутые косы продеваем ленту и завязываем бант. Прическа готова. Конечно, сразу плетение не получится идеальным. Не расстраивайтесь и продолжайте тренироваться. Можно попробовать заплести рыбий хвост подруге или сестре. Попрактикуйтесь сначала на чужих волосах. Когда руки освоят новый вид плетения, и вы сможете заплетать эту косу, как говорится, «не глядя», то тогда вновь вернетесь к плетению на своих волосах. В конце концов, ваш колосок станет красивым и обворожительным.
Расчесываем волосы на две стороны. На каждой стороне должно быть одинаковое количество волос. Работаем с левой стороны. У виска от пробора отделяем тонкую прядь волос. Перекручиваем ее в сторону пробора. Держим жгут правой рукой. Левой рукой отделяем другую прядь. Она должна быть точно под первой. Делаем жгут из новой пряди. Переплетаем два жгута между собой, первый под вторым. Перекладываем оба жгута в правую руку и смешиваем в одну прядь. Повторяем пункты 4-8. Закрепляем концы волос под косой с помощью шпилек. Переходим к правой половине волос и повторяем все с начала. Под подогнутые косы продеваем ленту и завязываем бант. Прическа готова. Конечно, сразу плетение не получится идеальным. Не расстраивайтесь и продолжайте тренироваться. Можно попробовать заплести рыбий хвост подруге или сестре. Попрактикуйтесь сначала на чужих волосах. Когда руки освоят новый вид плетения, и вы сможете заплетать эту косу, как говорится, «не глядя», то тогда вновь вернетесь к плетению на своих волосах. В конце концов, ваш колосок станет красивым и обворожительным.
способы создания модных причесок Прическа с колоском и кудрями
Аккуратная и стильная прическа — это визитная карточка каждой женщины. Если вы счастливая обладательница длинной и пышной шевелюры, которую необходимо быстро, красиво и качественно преобразить в шикарную прическу, мы с удовольствием расскажем вам, как плести колосок. Специально для этого красивого занятия мы собрали множество видео уроков и мастер-классов с фото для начинающих , которые помогут вам создавать стильные прически из кос как на праздник , так и на каждый день .
ак заплести колосок: простая техника для начинающих
Прическа, получившая в народе название «колосок», также
именуется
«дракончик» или «французская коса». Это
не просто красивый, а и практичный
способ,
ведь
он помогает привести в порядок непослушные пряди, убрать их подальше от лица.
Данная косичка является классическим вариантом плетения, на основе которого можно создать множество других прекрасных причесок. Вот почему
так важно поскорее узнать, как заплести колосок,
и мы предлагаем немедленно приступить к этому занятию.
Вот почему
так важно поскорее узнать, как заплести колосок,
и мы предлагаем немедленно приступить к этому занятию.
1. Чтобы вам было проще сделать свою первую французскую косичку (она же колосок), слегка намочите волосы или нанесите мусс. Тщательно расчешите волосы. Также рекомендуем вам собрать волосы от макушки с помощью силиконовой резинки. Так прическа будет лучше зафиксирована, и с ней легче управиться.
2. Волосы, собранные в хвостик, послужат в качестве центральной пряди. По бокам отделяем еще по одной пряди аналогичного размера.
3. Прядь, которая находится справа, мы переносим в центр.
4. Туда же мы перекладываем и левую прядку.
5. Далее мы будем добавлять пряди волос из оставшейся шевелюры и аккуратно вплетать их в колосок. Подбираем прядку справа и накладываем ее на прядь, оказавшуюся на данный момент с правой стороны (прядь №2). Далее переносим эту двойную прядь через верх в середину.
6. А
налогичным образом поступаем с левой стороны.
А
налогичным образом поступаем с левой стороны.
7. Чередуем плетение, прибавляя пряди поочередно с разных сторон.
8. Доходи до области шеи и получаем аккуратно уложенные пряди. Продолжаем плести, как обычную косу.
Вот ваш первый колосок и подошел к концу! Однако, на самом деле, это начало ваших новых причесок и завораживающих образов. Вы можете немного расправить косичку , получив объемный вариант прически . Также попробуйте заплести сразу два колоска, предварительно разделив свои волосы на 2 равные части. Готовую прическу можно зафиксировать лаком, а резинку, которую мы использовали в самом начале, аккуратно спрятать под заколкой или обрезать при помощи ножниц.
Е
ще одно плетение, аналогичное предыдущему варианту, называется «французская коса навыворот».
Для того чтобы правильно научиться плести этот колосок, внимательно рассмотрите фотоинструкцию
.
Хорошо расчешите волосы и отберите часть волос слева или справа от пробора. Разделите волосы на 3 равных части.
Разделите волосы на 3 равных части.
Прядь волос, которая находится справа, заводим под центральную прядь.
Прядь, находящуюся слева, заводим, и она оказывается в центре.
Теперь мы будем подхватывать пряди поочередно с двух сторон. Отделяем справа, из общей массы волос тоненькую прядку и присоединяем ее к пряди, находящейся в плетении справа.
Эту двойную прядь мы заводим под центральную прядь.
С другой стороны аналогично: отделяем тонкую прядку слева и соединяем ее с левой прядью из плетения. Обе эти пряди отправляем под среднюю через низ.
Продолжаем плести, пока не закончатся пряди для подхвата. Далее вяжем хвостик или обычную косу, продолжая укладывать пряди под низ. Если вы начнете плетение над ухом, то ваша косичка будет постепенно спадать на бок. Расправьте ее, так плетение покажется более объемным и вы сможете применять его даже на средние волосы.
Многих девушек интересует вопрос, как заплести колосок самой себе на длинные волосы . Вы повторяете те же действия, что были описаны выше. Рассмотрите еще раз внимательно схему и фотоинструкцию.
Вы повторяете те же действия, что были описаны выше. Рассмотрите еще раз внимательно схему и фотоинструкцию.
На короткие волосы по тому же принципу вы можете сделать очень красивую прическу из двух французских косичек по бокам. Так ие косы мы плетем до затылка наоборот или наизнанку, а затем закрепляем по центру невидимками.
Красивые прически самой себе вы можете заплести по следующим схемам.
Колосок с хвостиком самой себе
Коса со жгутами — оригинальная прическа для повседневного выхода. Праздничная прическа своими руками — это возможно.
Как заплести рыбий хвостик самой себе?
А вот еще одна дивная прическа — рыбий хвост . Обучение этой технике плетения мы также предлагаем по схеме.
- Расчешите волосы и разделите на 2 части.
- Слева отделяем тонкую прядь волос и отправляем ее в правую часть.
- Точно так же перебрасываем справа налево прядь аналогичного размера. Пряди должны быть одинаковыми, чтобы прическа получилась аккуратной.
- Плетем по указанной схеме до конца и закрепляем волосы резинкой.
- Такая прическа как рыбий хвостик, очень красиво смотрится на ровных, длинных волосах. Чем больше волос, тем более объемный выходит рыбий хвост.

Как заплести колосок ребенку: пошаговая инструкция
Маленькие модницы также обожают красивые и стильные прически. Однако они часто не могут и пяти минут усидеть на месте и требуют мгновенного результата. Поэтому мы разработали шуточный алгоритм маминых действий для эффективного плетения косичек и колосков.
Итак, наш вариант действий, как заплести колосок ребенку — пошаговая инструкция к вашим услугам.
Вариант 1
- Берете ребенка (девочку с длинными или средними волосами).
- Усаживаете на стул.
- Даете в руки планшет с играми.
Вариант 2 мало чем отличается от предыдущего:
- Девочка.
- Стул.
- Включаем мультики.
- Показываем готовый результат.

Ранее мы писали, какие чудесные можно заплести своими руками. А вот и схемы плетения с фото инструкцией для мамочек, желающих сделать красивую прическу своей дочурке.
Объемный колосок на бок , начиная со лба. Плетем по схеме, которую мы уже не раз проходили, добавляя с боку прядки и скручивая их в виде жгутов.
«Колосок», «дракончик», «французская коса», «рыбий хвост» — все эти названия носит классическая женская прическа, отличающаяся красотой, удобством и простотой в плетении. Это один из способов заплетать косичку, подходящей для женщин любого возраста. А если научиться основному варианту колоска, то, отталкиваясь от этого, можно варьировать прическу различным образом.
Французская коса — не новое изобретение. Появилась она — как мы поняли из названия — во Франции, а позже то выходила из моды, то возвращалась. Но есть пожалуй веские основания считать этот вид прически классикой, ведь это и красиво, и элегантно, и удобно… Такие прически не должны исчезать из применения: простота и красота — вечны!
Но лет 10 назад подход и взгляд на колосок был другим. Тогда считалось, что герой нашего повествования подходит только для торжественных событий, для элегантного и эффектного выхода. Прикольный и игривый рыбий хвостик был серьезной, строгой прической.
Тогда считалось, что герой нашего повествования подходит только для торжественных событий, для элегантного и эффектного выхода. Прикольный и игривый рыбий хвостик был серьезной, строгой прической.
Сейчас нет такого ограниченного подхода. Колосок подходит практически для любого повода: дружеская встреча, вечеринка, деловая поездка, прогулка… да как хотите. Единственный критерий — правильное плетение. Вот здесь действительно надо усвоить главные уроки.
Базовая техника плетения колоска
Итак, существует довольно простая схема, которая позволит легко заплести колосок своими руками. Мы проанализируем разные варианты, но основной вид косы, на котором мы будем основываться следующий:
Выполняйте каждый пункт из пошаговой инструкции. Мы будем поэтапно объяснять, чтобы вы сумели правильно уложить волосы, как на рисунках.
- Намочите немного свои волосы — совсем чуть-чуть. Это сделает их более послушными и позволит укладывать аккуратнее. Если есть мусс, то можно воспользоваться им. Главное, чтобы волосы стали послушнее и не расплетались. Соберите прядь волос на макушке, как показано на рисунке 3. Завяжите резинкой, так как во время плетения обе руки у нас будут заняты, а прядей будет три.
Рисунок 3. Резинка удерживает первую прядь волос - Оставив в центре зафиксированную прядь, соберите еще две по обе стороны от нее.
- Переложите правую прядь в левую сторону, а центральную перенесите вправо так, чтобы она пролегала под правой прядью волос.
- Уложите левую прядь также вправо, но она должна укладываться над правой, как показано на рисунке.
- Теперь проведите над левой прядью, которая в данный момент находится с правой стороны, нашу центральную прядь, но обязательно сплетите с ней вместе дополнительную, предварительно собрав в один пучок. На голове уже должен формироваться объемный узел основания колоска.
- Продолжайте по той же схеме с обеих сторон, пока не дойдете до области шеи.
- Дойдя до области шеи, сделайте продолжение в виде обычной косы. Таким образом хвост колоска будет закончен.
 Главное, чтобы волосы стали послушнее и не расплетались. Соберите прядь волос на макушке, как показано на рисунке 3. Завяжите резинкой, так как во время плетения обе руки у нас будут заняты, а прядей будет три.
Главное, чтобы волосы стали послушнее и не расплетались. Соберите прядь волос на макушке, как показано на рисунке 3. Завяжите резинкой, так как во время плетения обе руки у нас будут заняты, а прядей будет три. Ну вот, колосок готов. Резинку, придерживающую центральную прядь либо аккуратно обрежьте и снимите, либо спрячьте под прической. Нанесите немного лака и чуть-чуть расправьте, чтобы дракончик приобрел объемный вид.
Данная пошаговая инструкция — лишь база, простейший фундамент, на котором основано множество других видов колоска. Теперь, обладая знаниями об этой базе, вы сможете изменять вид вашей прически сообразно своим фантазиям.
Французская косичка, заплетаемая на бок
Красиво и аккуратно будет смотреться, например, колосок, заплетаемый на бок. Это и стильно, и молодежно, неплохо смотрится и на голове взрослых женщин. В принципе, схема та же самая, как мы и говорили, только, добавляя боковые пряди, необходимо перекручивать ее как жгут.
Шаг за шагом:
- Расчешите волосы назад;
- С левой или правой стороны соберите прядь побольше и сделайте из нее три небольшие пряди;
- Начните делать обычный колосок, как на первых шагах нашей инструкции — до того, как мы начали вплетать дополнительные пряди;
Вплетите в косичку дополнительную небольшую прядь с той стороны, которую выбрали в первом пункте данной инструкции; - Добавьте следующую прядь с другой стороны головы;
- По очереди добавляйте пряди с той и другой стороны, не забывая при этом затягивать косичку, чтобы волосы не расплетались и колосок не терял своей формы;
- В финальной части все — то же самое, что и в обычном колоске. У основания шеи заплетите обычную косу. Закрепите шпильками звенья косы и аккуратно расправьте каждую, чтобы прическа стала объемней.
На рисунке 10 вы можете рассмотреть поэтапную иллюстрацию колоска на бок, чтобы проследить за выполнением всех пунктов инструкции правильно.
Рисунок 10. Косичка на бок
Французская косичка навыворот
Этот вид косички мало чем отличается от классического колоска по способу своего плетения. Выглядит он очень солидно, звенья отличаются выраженностью и пухлостью. Несомненно, такая прическа будет выглядеть весьма элегантно.
Собственно, теперь вы умеете плести уже несколько видов французской косички. Проявляйте фантазию, вы можете использовать совершенно разные способы для того, чтобы колосок смотрелся эффектней, красивее. Регулируйте тугость косичек, чтобы они не спадали и не портились, а с другой стороны, не были затянуты чересчур сильно. Меняйте центральную косичку, чтобы смещать расположение всей косы. Экспериментируйте с цветом и используйте лак для волос.
Можно заплетать колосок вокруг головы или делать во французском стиле. Вот, как выглядит последний:
Эту же прическу можно сделать сдвоенной. Кстати, отличное решение для вашей доченьки.
Техника плетения колоска, в основном, подчиняется одним и тем же законам, как вы могли видеть. Удивительно, но это по-настоящему универсальная прическа, при это всегда выглядит красиво и аккуратно. Дракончик не позволяет волосам падать на лицо, что очень удобно и практично.
Заплетая косички, старайтесь захватывать пряди с каждой из сторон в одинаковом количестве, равномерно. От того, сколько волос вы захватите, будет зависеть форма, а значит красота вашей прически. Кроме того, надо сказать, что французская косичка плетется, как правило, для прямых волос, поэтому если у вас кудри, то придется основательно погладить их утюжком, выровнять, чтобы внешний вид колоска не испортился.
Можно по-разному украшать косичку: стразами, бантиком, ободком, различными заколками. В этом, пожалуй, дело ограничивается только вашим вкусом.
Если хотите, то сделайте два колоска по бокам. Не обязательно заплетать его туго и собранно. Опустите волоски, расправьте прическу, чтобы она выглядела так, будто носите вы ее не первый день. Посмотрите на рисунок 21 и убедитесь сами:
Рисунок 21. Добавьте простоты
Видите — нет необходимости создавать строгую прическу. Импровизируйте.
Не помешает выпустить несколько прядей из косы. Опустите их так, чтобы они небрежно спадали с обеих сторон головы.
Если вы сделали два колоска, то разве есть необходимость приводить их в зеркально-симметричный вид? Зачем? Ведь прическа будет интересно выглядеть, если они будут абсолютно ассиметричны. Отпустите вожжи и отбросьте шоры! — позвольте своей фантазии разыграться. Поверьте, этот совет не менее важен, чем пошаговая инструкция!
Колосок занимает одну из лидирующих позиций среди причесок. Он стал столь популярным из-за оригинальности плетения. Кроме того, коса красиво подчеркивает изгибы головы девушки, густые волосы.
Эту прическу можно носить на любом мероприятии, ведь колосок универсален и уместен всегда. Если хочется заплести его для торжественного события, добавьте шпильки с жемчужинами, ленты, заколку с искусственными цветами. Для похода на учебу ограничьтесь простой резинкой.
Схема лёгкий
простой на долго
стильно модно причёска
За несколько лет своего существования коса несколько раз видоизменялась и принимала разнообразные формы. Сегодня она получила полную свободу плетения. На фото с прической видно, что колосок может быть заплетен различными способами. Все зависит от вашей фантазии.
Современная коса дополняется веяниями моды. Раньше она была тугой, гладкой, а сейчас стала более небрежной и свободной.. Особенно красиво колосок смотрится на брондированной или колорированной шевелюре.
Плетение колоском хорошо подойдет для маленьких девочек, юных леди и взрослых женщин. Прическа простая, модная и устойчивая.
Конечно же, несколько дней подряд проносить ее нельзя. Но, если заплести косу с утра и сбрызнуть лаком сильной фиксации, укладка спокойно продержаться до вечера.
Давайте разберёмся с тем, как правильно плести популярную прическу колосок, и узнаем несколько секретов.
Популярная коса идеально выглядит на волосах любой длины. При большом старании, его получится сделать и на короткой шевелюре, если, конечно же, вы не стрижены под мальчишку.
За счет различных вариантов косы, она идеально дополнит ваш образ, добавит ему женственности, изящности. Колосок отлично смотрится при любой форме лица.
Довольно часто складывается такая ситуация, что при плетении косы волосы выскакивают, поэтому появляются «петухи». Часто это происходит при попытке соорудить прическу в виде 2 колосков.
У опытных парикмахеров есть секреты которые облегчают процесс и делают косу аккуратной. Посмотрите фото модной прически колосок на длинные или средние волосы, вооружитесь расческой, заколками. При должной сноровке вам потребуется максимум 10 минут.
- изящная и аккуратная косичка получается из тоненьких прядей. Когда хочется объемный вариант, возьмите крупные пряди.
- для создания небрежной, слегка свободной косы, следует брать большие пряди волосы и перекрещивать их реже.
- расчёсывайте и подтягивайте каждую прядку, чтобы избежать появления «петухов».
- волосы часто рассыпаются во время плетения, что усложняет процесс. Слегка сбрызните их лаком или обычной проточной водой. Также можно смочить руки.
- берите прядки одинаковой толщины. В этом случае коса не будет заворачиваться на бок.
- если хочется, чтобы прическа была объемнее и пышнее, потяните пальцами некоторые звенья косички. Но делайте это аккуратно, чтобы она не рассыпалась.
- небольшие «петухи» устраняются расческой с мелкими зубьями.
- если вы планируете соорудить косу вокруг головы, для начала возьмите мелкий гребешок и сделайте небольшой начес у корней. Это позволит укладке выглядеть более элегантной.
Также просмотрите фото .
Не переживайте, если с первого раза у вас не получится идеальная прическа. Прежде чем заплетать ее себе, потренируйтесь на другом человеке. Руки должны привыкнуть держать несколько прядей одновременно.
Колосок хорошо подходит еще и в тех случаях, когда традиционный хвостик уже надоел, а волосы нужно забрать. Например, вовремя похода в бассейн, при ветреной или дождевой погоде. Украсьте прическу различными аксессуарами: ободком, шпильками, яркими заколками.
Инструкция по созданию прически
Существует масса вариантов колосков: на выворот, на бок, вокруг головы, детские варианты с бантиками, красивые прически в виде двух колосков. Но сначала стоит научиться плести классический колосок. Различные видео с этой прической помогут овладеть техникой.
- Причешите волосы, отделите небольшой локон у основания волос. Выбранную прядь разделите на три части.
- Перекрестите пряди, затем перебросьте через одну из крайних дополнительный боковой локон, находящийся на той же стороне. Включите его в центр плетения. Боковые прядки поочередно добавляйте к плетению.
- Плетите косу аналогичным образом, пока не добавите в нее последние боковые прядки.
Необычно смотрится вывернутый колосок. Соорудить его не намного сложнее классического. Достаточно один раз посмотреть видео, в котором рассказывается, как плести эту прическу. Это более эффектный, выразительный вариант.
- Расчешите шевелюру, на макушке головы отделите центральную, левую, правую пряди.
- Левую прядь положите под центральную, а правую – под центральную и левую.
- Затем возьмите центральный локон, присоедините к нему свободные пряди и подложите под правый. Обогните его таким образом, чтобы он оказался в центре косы.
- Аналогичным образом заплетите левую прядку. Также захватите часть свободных волос, но левая прядь должна оставаться в центре косы.
- Вывернутую косу продолжайте плести до тех пор, пока не закончатся все волосы.
- Кончик косички закрепите резинкой или заколкой.
Еще несколько вариантов
Если на праздник хочется сделать необычную прическу из привычных колосков, попробуйте заплести их вокруг головы. Здесь нет ничего сложного. Главное, найти картинки с изображением прически, а подробная схема подскажет порядок действий.
- Волосы расчешите, слегка увлажните.
- По центру головы сделайте ровный пробор, разделяющий пряди на две ровные части.
- За правым виском отделите три локона, как для обычной косички, а затем начинайте плетение по спирали. С каждым последующим шагом добавляйте новые прядки.
- Плетение заканчивается около левого виска. Кончик косички спрячьте под волосы, закрепите заколкой невидимкой.
- Таким же образом плетется коса от левого виска. Можно сделать наоборот, и начать плетение с правой стороны. При желании прикрепите по бокам заколочки.
Теперь взгляните на и .
Прическе отведена особая роль в образе любой женщины, ведь окружающим бросаются в глаза, в первую очередь, и ногти. Умение красиво укладывать шевелюру не только помогает дамам самостоятельно создавать очаровательные образы, но и существенно экономит семейный бюджет, так как отпадает необходимость часто посещать парикмахера. Например, колосок – несложная прическа и освоив основные принципы плетения, женщина с легкостью сможет сама себе его заплетать.
Прическа колосок (сложная)
Колосок прекрасно подходит для волос любой длины. Стильная коса послужит украшением как молодым , так и женщинам постарше.
Вследствие того, что современные виды этой прически имеют самые разнообразные модификации, она отлично впишется в созданный образ и придаст особый шарм и женственность представительницам прекрасного пола.
Колосок – универсальная прическа. Особенность ее заключается в том, что она смотрится с любой формой лица. Какое бы плетение не было выбрано – воздушное или гладкое, в любом случае, необычная косичка придаст нежность и привлекательность образу. А колосок, выполненный оригинальным способом, привлечет к вашей прическе взоры окружающих.
Чтобы процесс плетения максимально облегчить, но в итоге получить красивый и аккуратный колосок, нужно следовать некоторым простым правилам:
- Изящная косичка получается из тонких прядей.
- Чтобы придать косам объем, лучше использовать крупные пряди.
- Для получения , небрежной, слегка распущенной косы, локоны берутся большими, а их перекрещивание следует делать пореже.
- Плести станет значительно проще, а волосы не будут рассыпаться в том случае, если их немножко обрызгать водой или каким-нибудь средством (муссом, гелем).
- Чтобы не появлялись «петухи», каждую новую прядку рекомендуется расчесывать и подтягивать.
- Во избежание заворачивания косички набок, нужно делать одинаковой толщины.
- Для избавления от мелких «петухов» используется расческа с маленькими зубчиками.
- Колосок, выполненный во французском стиле, будет смотреться пышнее и объемнее, если некоторые из его звеньев слегка потянуть пальцами.
- Для создания оригинального варианта колоска можно добавлять в плетение различные аксессуары – ленты, ободки, стразы, заколки.
Любая прическа с первого раза не может получиться идеальной, поэтому, прежде, чем делать ее себе, лучше потренироваться на ком-то еще или на манекене.
Учимся плести классический колосок
Многих женщин интересует, как научиться . На сегодняшний день существует достаточно много видов и способов плетения этой косички.
Одним из самых популярных считается классический колосок. Для того, чтобы в итоге получилась красивая и аккуратная прическа, необходимо использовать специальные укладочные средства:
- Пенку или мусс для волос
- расческу с тонким хвостиком
- резинку
Этапы плетения классического колоска:
- Для начала волосы тщательно расчесываются, в этом случае отделение прядей будет проходить без труда.
- С того места, откуда планируете начинать плетение, нужно от остальных волос отделить небольшую прядь.
- Этот локон делится на три части, которые один раз между собой перекручиваются. Сначала правая прядь меняется местами со средней, после этого левая меняется с той, которая посередине.
- Большим пальцем от общей пряди волос отделяется новый локон, который добавляется к правой части.
- Перекручиваем ее со средней.
- После этого новая прядь добавляется к левой и снова перекручивается со средней.
- Подобная схема плетения продолжается, новые пряди поочередно добавляются то к правой, то к левой, пока не будут использованы все волосы.
- Доплетаем колосок как обычную косичку.
Как плести колосок расскажет видеоматериал:
Создаем прическу «2 колоска»
Прическа «2 колоска»
Для тех, кто умеет заплетать один колосок, не составит труда сделать прическу в 2 колоска. Для создания подобного шедевра, важно сделать идеально ровный пробор.
Какой из многочисленных видов колоска вы выберите, зависит исключительно от вас. Все способы будут смотреться и изящно.
- Подготовка к плетению заключается в тщательном расчесывании волос.
- После этого локоны делятся на прямой пробор.
- Сначала колосок заплетается по одну сторону от пробора, потом по другую.
- Около челки, с левой стороны от пробора, захватывается прядь волос, которую следует разделить на три одинаковые части.
- Далее, техника плетения зависит от вида косы.
- Готовая прическа красиво укладывается и украшается. Можно косички оформить в виде спиральки или колечка.
- Первый этап аналогичен созданию варианта в классическом стиле. Отделенная на макушке прядь волос распределяется на три равные части.
- Но после этого левую прядь следует поместить не на центральную, а под нее.
- Правую прядь аккуратно заносим под левую и центральную.
- Следующим этапом к средней пряди добавляем часть свободных волос и все это заводим под правую. При этом ее нужно обогнуть таким образом, чтобы она вновь появилась в центре косички.
- Аналогично поступаем с левой прядью. Но теперь посередине должна оказаться именно она.
- Вывернутый колосок заплетается, пока есть свободные волосы.
- В конце скрепить резинкой.
Колосок «Вокруг головы»
Такой вариант плетения является не совсем стандартным, но выглядит на голове великолепно.
Особенно эффектно такая прическа будет смотреться на густой . Для тонких волос имеет смысл у корней сделать легкий начес.
- Перед плетение пряди следует хорошенько расчесать и увлажнить.
- Посередине головы делается ровный пробор, чтобы волосы получились разделенными на две ровные половинки.
- За правым виском выделяются три одинаковые прядки, так же, как и при классическом варианте.
- Косичку следует начинать плести по спирали, каждый раз добавляя новые пряди. В работе должны быть задействованы все волосы.
- Завершается плетение с противоположной стороны, у левого уха.
- Кончик косички лучше спрятать под локоны и закрепить шпилькой или невидимкой, чтобы он не выглядывал и не вылезал.
- Подобным способом начинаем плести полукругом от левого виска.
- Кончик волос также нужно спрятать под звенья косы и зафиксировать при помощи невидимок.
- Оригинально будет смотреться такая прическа, если украсить ее заколками.
Плести косу вокруг головы сложнее, чем классический вариант, поэтому для начала желательно потренироваться на ком-то.
Колосок «На бок»
Весьма интересен вариант колоска, заплетенного на бок. Техника создания по принципу схожа с классическим вариантом, за тем лишь исключением, что каждая прядь, добавляемая сбоку, перекручивается в виде жгутика.
- Первый этап аналогичен предыдущим – локоны тщательно расчесываются.
- Затем с правой или левой стороны (неважно с какой), отделяется небольшой локон средней толщины и делится на три части.
- Первые несколько звеньев плетутся по технологии классической косы.
- Далее в нашу косичку добавляется локон, взятый с любой из сторон. С этого момента начинается уже плетение косы на бок.
- Следом отделяем прядь с противоположной стороны и также добавляем ее в плетение.
- Продолжаем поочередно с каждой стороны добавлять в колосок новые прядки, стараясь подтянуть потуже.
- Уже от основания шеи продолжаем заплетать как обычную косичку.
- На конце закрепляем все резинкой.
- Украшение готовой прически каждый выполняет по своему усмотрению.
Особенно популярен в последнее время зигзагообразный колосок. Создание необычной прически сводится к следующим действиям:
- Плетение рекомендуется начинать чуть ниже левого виска. Как только задействованные пряди оказываются отделенными, начинается плетение в противоположном направлении. Причем, локоны в косичку добавляются исключительно с левой стороны.
- После того, как будут готовы 5-6 плетений, колосок разворачивается влево.
- Продолжаем плести дальше, только теперь прядки добавляются только с правой стороны. Длина подобного плетения равна длине первого элемента.
- Таким образом зигзаг доводится до затылочной области, где и фиксируется при помощи обычной заколки.
Двойной колосок – еще один достойный вид популярной в последнее время прически. Процедура создания прически не совсем проста.
- На первом этапе плетение осуществляется как при вывернутом колоске, каждые следующие прядки подкладываются друг под друга.
- Далее к крайним локонам добавляются дополнительные волосы, отделенные горизонтальным пробором. Такие подхваты следует добавлять к крайним прядям поочередно, то к левой, то к правой.
- После этого от основного локона отделяется незначительная часть, которая сверху закрепляется зажимом.
- Делается последующий подхват, и косичка сплетается под низ. Аналогичная процедура проводится и с другой стороны.
- Когда часть колоска уже готова, необходимо потихоньку вытянуть локоны за края. Заниматься этим потом нежелательно. Мало того, что сделать это окажется намного труднее, так еще и коса потеряет свою опрятность и привлекательность.
- После этого следует продолжать привычное плетение.
- После того, как свободные пряди окажутся вплетенными, нужно сделать основное плетение посредством выделения и закалывания небольших , но уже без подхватов.
- Готовый колосок необходимо зафиксировать внизу при помощи зажимов.
- Для создания верхнего колоска необходимо взять свободную прядь, расположенную сверху, разделить ее на три части и начинать плетение обычной косы. Локоны при этом должны накладываться поверх, друг на друга.
- Как и при оформлении колоска нужно выполнять подхват из следующих свободных прядей, поделенных на части.
- Плетение продолжается до конца.
- В завершении обе косы фиксируются резинками.
Колосок на короткие волосы
Колосок превосходно смотрится и на коротких волосах. Один из вариантов предполагает следующие действия:
- Для начала волосы нужно отделить пробором через темечко, от верхней части одного уха до другого.
- Плести следует начинать от самого уха, постепенно добавляя локоны от линии волос на виске.
- Кончики прически скорее всего будут торчать из косы. Чтобы созданному шедевру придать аккуратности, необходимо воспользоваться гелем для волос, пригладив проблемные участки или просто заколоть их маленькими шпильками.
Современные модные тенденции вдохнули новую жизнь в забытый колосок. Научившись плести очаровательную косичку, милые дамы смогут самостоятельно экспериментировать с расположением плетения, радуя окружающих все новыми .
Вконтакте
Плетение 2 колосков пошаговая инструкция. Пошаговая инструкция выполнения
“Колосок”, “дракончик”, “французская коса”, “рыбий хвост” – все эти названия носит классическая женская прическа, отличающаяся красотой, удобством и простотой в плетении. Это один из способов заплетать косичку, подходящей для женщин любого возраста. А если научиться основному варианту колоска, то, отталкиваясь от этого, можно варьировать прическу различным образом.
Французская коса – не новое изобретение. Появилась она – как мы поняли из названия – во Франции, а позже то выходила из моды, то возвращалась. Но есть пожалуй веские основания считать этот вид прически классикой, ведь это и красиво, и элегантно, и удобно… Такие прически не должны исчезать из применения: простота и красота – вечны!
Но лет 10 назад подход и взгляд на колосок был другим. Тогда считалось, что герой нашего повествования подходит только для торжественных событий, для элегантного и эффектного выхода. Прикольный и игривый рыбий хвостик был серьезной, строгой прической.
Сейчас нет такого ограниченного подхода. Колосок подходит практически для любого повода: дружеская встреча, вечеринка, деловая поездка, прогулка… да как хотите. Единственный критерий – правильное плетение. Вот здесь действительно надо усвоить главные уроки.
Базовая техника плетения колоска
Итак, существует довольно простая схема, которая позволит легко заплести колосок своими руками. Мы проанализируем разные варианты, но основной вид косы, на котором мы будем основываться следующий:
Выполняйте каждый пункт из пошаговой инструкции. Мы будем поэтапно объяснять, чтобы вы сумели правильно уложить волосы, как на рисунках.
- Намочите немного свои волосы – совсем чуть-чуть. Это сделает их более послушными и позволит укладывать аккуратнее. Если есть мусс, то можно воспользоваться им. Главное, чтобы волосы стали послушнее и не расплетались. Соберите прядь волос на макушке, как показано на рисунке 3. Завяжите резинкой, так как во время плетения обе руки у нас будут заняты, а прядей будет три.
Рисунок 3. Резинка удерживает первую прядь волос - Оставив в центре зафиксированную прядь, соберите еще две по обе стороны от нее.
- Переложите правую прядь в левую сторону, а центральную перенесите вправо так, чтобы она пролегала под правой прядью волос.
- Уложите левую прядь также вправо, но она должна укладываться над правой, как показано на рисунке.
- Теперь проведите над левой прядью, которая в данный момент находится с правой стороны, нашу центральную прядь, но обязательно сплетите с ней вместе дополнительную, предварительно собрав в один пучок. На голове уже должен формироваться объемный узел основания колоска.
- Продолжайте по той же схеме с обеих сторон, пока не дойдете до области шеи.
- Дойдя до области шеи, сделайте продолжение в виде обычной косы. Таким образом хвост колоска будет закончен.
Ну вот, колосок готов. Резинку, придерживающую центральную прядь либо аккуратно обрежьте и снимите, либо спрячьте под прической. Нанесите немного лака и чуть-чуть расправьте, чтобы дракончик приобрел объемный вид.
Данная пошаговая инструкция – лишь база, простейший фундамент, на котором основано множество других видов колоска. Теперь, обладая знаниями об этой базе, вы сможете изменять вид вашей прически сообразно своим фантазиям.
Французская косичка, заплетаемая на бок
Красиво и аккуратно будет смотреться, например, колосок, заплетаемый на бок. Это и стильно, и молодежно, неплохо смотрится и на голове взрослых женщин. В принципе, схема та же самая, как мы и говорили, только, добавляя боковые пряди, необходимо перекручивать ее как жгут.
Шаг за шагом:
- Расчешите волосы назад;
- С левой или правой стороны соберите прядь побольше и сделайте из нее три небольшие пряди;
- Начните делать обычный колосок, как на первых шагах нашей инструкции – до того, как мы начали вплетать дополнительные пряди;
Вплетите в косичку дополнительную небольшую прядь с той стороны, которую выбрали в первом пункте данной инструкции; - Добавьте следующую прядь с другой стороны головы;
- По очереди добавляйте пряди с той и другой стороны, не забывая при этом затягивать косичку, чтобы волосы не расплетались и колосок не терял своей формы;
- В финальной части все – то же самое, что и в обычном колоске. У основания шеи заплетите обычную косу. Закрепите шпильками звенья косы и аккуратно расправьте каждую, чтобы прическа стала объемней.
На рисунке 10 вы можете рассмотреть поэтапную иллюстрацию колоска на бок, чтобы проследить за выполнением всех пунктов инструкции правильно.
Рисунок 10. Косичка на бок
Французская косичка навыворот
Этот вид косички мало чем отличается от классического колоска по способу своего плетения. Выглядит он очень солидно, звенья отличаются выраженностью и пухлостью. Несомненно, такая прическа будет выглядеть весьма элегантно.
- С левой или правой стороны от цезуры волос выберите прядь побольше и разделите ее на три равные прядки.
- Правую прядку переложите под центральной. А последнюю перенесите вверх и вправо как на рисунке 12.
Рисунок 12. Первое звено - Затем прядку слева проносите под правой и она выходит в центральную часть косички.
- Начинайте подхватывать пряди с двух сторон косички, как уже многократно было показано.
- Соедините дополнительную прядь с центральной, которая находится теперь справа от косички.
- Две пряди, соединенные вместе, проведите под центральной.
- Ровно то же самое сделайте с левой стороной.
- Продолжайте плести звенья колоска до тех пор, пока не закончатся дополнительные прядки. Затем сплетите обычную косу, завершив прическу хвостиком. Расправьте, чтобы появился объем.
Собственно, теперь вы умеете плести уже несколько видов французской косички. Проявляйте фантазию, вы можете использовать совершенно разные способы для того, чтобы колосок смотрелся эффектней, красивее. Регулируйте тугость косичек, чтобы они не спадали и не портились, а с другой стороны, не были затянуты чересчур сильно. Меняйте центральную косичку, чтобы смещать расположение всей косы. Экспериментируйте с цветом и используйте лак для волос.
Можно заплетать колосок вокруг головы или делать во французском стиле. Вот, как выглядит последний:
Эту же прическу можно сделать сдвоенной. Кстати, отличное решение для вашей доченьки.
Техника плетения колоска, в основном, подчиняется одним и тем же законам, как вы могли видеть. Удивительно, но это по-настоящему универсальная прическа, при это всегда выглядит красиво и аккуратно. Дракончик не позволяет волосам падать на лицо, что очень удобно и практично.
Заплетая косички, старайтесь захватывать пряди с каждой из сторон в одинаковом количестве, равномерно. От того, сколько волос вы захватите, будет зависеть форма, а значит красота вашей прически. Кроме того, надо сказать, что французская косичка плетется, как правило, для прямых волос, поэтому если у вас кудри, то придется основательно погладить их утюжком, выровнять, чтобы внешний вид колоска не испортился.
Можно по-разному украшать косичку: стразами, бантиком, ободком, различными заколками. В этом, пожалуй, дело ограничивается только вашим вкусом.
Если хотите, то сделайте два колоска по бокам. Не обязательно заплетать его туго и собранно. Опустите волоски, расправьте прическу, чтобы она выглядела так, будто носите вы ее не первый день. Посмотрите на рисунок 21 и убедитесь сами:
Рисунок 21. Добавьте простоты
Видите – нет необходимости создавать строгую прическу. Импровизируйте.
Не помешает выпустить несколько прядей из косы. Опустите их так, чтобы они небрежно спадали с обеих сторон головы.
Если вы сделали два колоска, то разве есть необходимость приводить их в зеркально-симметричный вид? Зачем? Ведь прическа будет интересно выглядеть, если они будут абсолютно ассиметричны. Отпустите вожжи и отбросьте шоры! – позвольте своей фантазии разыграться. Поверьте, этот совет не менее важен, чем пошаговая инструкция!
Сделайте колосок “вниз головой”, т.е., чтобы плетение шло снизу вверх. Только в этом случае не стоит делать хвост в виде обычной косы, а просто заплетите оставшиеся волосы, завяжите так, чтобы выглядело красиво.
На самом деле, французская косичка – это прическа, которая ДОЛЖНА быть свободна от ограничений. Однообразие противоречит самому смыслу женской красоты, ведь главная цель – приятно удивить. Взгляните на рисунки 22 и 23.
Рис. 22. Разный подход
Рис. 23. Ещё один вариант
Кажется, что это – две разные косички. Нет! – Это простой колосок, уложенный со вкусом, но разными способами. Модели и мировые знаменитости пользуются этой элегантной и простой прической себе на пользу. Сделайте и вы модную и красивую косичку себе, подруге или дочке. Можете быть уверенной, что при достаточной ловкости и старательности, получится просто великолепно.
Видео уроки по плетению колосков из волос
Колосок — причёска очень практичная и бесконечно разнообразная, а главное, придуманная для удобства женщины. Лет 20 назад французские косы считались отличительной чертой старшеклассниц. Сейчас же нет, коса вернулась в моду слегка в изменённом виде — чуть небрежная, свободная, с выпущенными прядками и очень женственная. При желании и наличии должного времени на тренировки можно научиться плести колосок самостоятельно.
Что собой представляет причёска
Колосок, он же французский колосок или французская коса — общее название для такого способа плетения волос, когда пряди захватываются от самой макушки, от пробора, а к ним приплетают волосы с боков — и так по всей длине. Свободные пряди собирают в так называемую русскую (обычную) косу и закрепляют резинкой или заколкой. Разновидностей колоска существует около десятка — это не считая тех причёсок, когда отдельными заплетёнными во французскую косу прядями оформляют распущенные волосы, чёлку или любую другую причёску, как конский хвост, например.
Французской косой или колоском можно разнообразить другие причёски
Как будет смотреться готовая причёска, зависит:
- от того, какая выбрана форма пробора;
- от количества прядей, их может быть две, три или четыре;
- от толщины прядей — в косу можно вплести только несколько тонких прядей или все;
- от направления плетения.
Заплестись можно таким образом:
На короткие волосы тоже есть вариант — коса Бохо. Можно сделать одну или две, по всей длине волос или на часть длины — от пробора и до уха. Плетение свободное, к тому же пряди можно чуть вытянуть, так что получится слегка небрежный и потому модный образ.
Чтобы научиться плести колосок, нужны правильные руки, врождённая аккуратность, немного терпения и подручные средства — зеркала, расчёска и резинки. Требуется немного, а вот навык плетения французской косы потом пригодится всему вашему окружению: детям, сёстрам, подругам и соседкам. Будете пользоваться спросом неимоверным, так что учитесь!
Аккуратная и стильная прическа — это визитная карточка каждой женщины. Если вы счастливая обладательница длинной и пышной шевелюры, которую необходимо быстро, красиво и качественно преобразить в шикарную прическу, мы с удовольствием расскажем вам, как плести колосок. Специально для этого красивого занятия мы собрали множество видео уроков и мастер-классов с фото для начинающих , которые помогут вам создавать стильные прически из кос как на праздник , так и на каждый день .
ак заплести колосок: простая техника для начинающих
Прическа, получившая в народе название «колосок», также именуется «дракончик» или «французская коса». Это не просто красивый, а и практичный способ, ведь он помогает привести в порядок непослушные пряди, убрать их подальше от лица. Данная косичка является классическим вариантом плетения, на основе которого можно создать множество других прекрасных причесок. Вот почему так важно поскорее узнать, как заплести колосок, и мы предлагаем немедленно приступить к этому занятию.
1. Чтобы вам было проще сделать свою первую французскую косичку (она же колосок), слегка намочите волосы или нанесите мусс. Тщательно расчешите волосы. Также рекомендуем вам собрать волосы от макушки с помощью силиконовой резинки. Так прическа будет лучше зафиксирована, и с ней легче управиться.
2. Волосы, собранные в хвостик, послужат в качестве центральной пряди. По бокам отделяем еще по одной пряди аналогичного размера.
3. Прядь, которая находится справа, мы переносим в центр.
4. Туда же мы перекладываем и левую прядку.
5. Далее мы будем добавлять пряди волос из оставшейся шевелюры и аккуратно вплетать их в колосок. Подбираем прядку справа и накладываем ее на прядь, оказавшуюся на данный момент с правой стороны (прядь №2). Далее переносим эту двойную прядь через верх в середину.
6. А налогичным образом поступаем с левой стороны.
7. Чередуем плетение, прибавляя пряди поочередно с разных сторон.
8. Доходи до области шеи и получаем аккуратно уложенные пряди. Продолжаем плести, как обычную косу.
Вот ваш первый колосок и подошел к концу! Однако, на самом деле, это начало ваших новых причесок и завораживающих образов. Вы можете немного расправить косичку , получив объемный вариант прически . Также попробуйте заплести сразу два колоска, предварительно разделив свои волосы на 2 равные части. Готовую прическу можно зафиксировать лаком, а резинку, которую мы использовали в самом начале, аккуратно спрятать под заколкой или обрезать при помощи ножниц.
Е
ще одно плетение, аналогичное предыдущему варианту, называется «французская коса навыворот».
Для того чтобы правильно научиться плести этот колосок, внимательно рассмотрите фотоинструкцию
.
Хорошо расчешите волосы и отберите часть волос слева или справа от пробора. Разделите волосы на 3 равных части.
Прядь волос, которая находится справа, заводим под центральную прядь.
Прядь, находящуюся слева, заводим, и она оказывается в центре.
Теперь мы будем подхватывать пряди поочередно с двух сторон. Отделяем справа, из общей массы волос тоненькую прядку и присоединяем ее к пряди, находящейся в плетении справа.
Эту двойную прядь мы заводим под центральную прядь.
С другой стороны аналогично: отделяем тонкую прядку слева и соединяем ее с левой прядью из плетения. Обе эти пряди отправляем под среднюю через низ.
Продолжаем плести, пока не закончатся пряди для подхвата. Далее вяжем хвостик или обычную косу, продолжая укладывать пряди под низ. Если вы начнете плетение над ухом, то ваша косичка будет постепенно спадать на бок. Расправьте ее, так плетение покажется более объемным и вы сможете применять его даже на средние волосы.
Многих девушек интересует вопрос, как заплести колосок самой себе на длинные волосы . Вы повторяете те же действия, что были описаны выше. Рассмотрите еще раз внимательно схему и фотоинструкцию.
На короткие волосы по тому же принципу вы можете сделать очень красивую прическу из двух французских косичек по бокам. Так ие косы мы плетем до затылка наоборот или наизнанку, а затем закрепляем по центру невидимками.
Красивые прически самой себе вы можете заплести по следующим схемам.
Колосок с хвостиком самой себе
Коса со жгутами — оригинальная прическа для повседневного выхода. Праздничная прическа своими руками — это возможно.
Как заплести рыбий хвостик самой себе?
А вот еще одна дивная прическа — рыбий хвост . Обучение этой технике плетения мы также предлагаем по схеме.
Расчешите волосы и разделите на 2 части.
Слева отделяем тонкую прядь волос и отправляем ее в правую часть.
Точно так же перебрасываем справа налево прядь аналогичного размера. Пряди должны быть одинаковыми, чтобы прическа получилась аккуратной.
Плетем по указанной схеме до конца и закрепляем волосы резинкой.
Такая прическа как рыбий хвостик, очень красиво смотрится на ровных, длинных волосах. Чем больше волос, тем более объемный выходит рыбий хвост.
Как заплести колосок ребенку: пошаговая инструкция
Маленькие модницы также обожают красивые и стильные прически. Однако они часто не могут и пяти минут усидеть на месте и требуют мгновенного результата. Поэтому мы разработали шуточный алгоритм маминых действий для эффективного плетения косичек и колосков.
Итак, наш вариант действий, как заплести колосок ребенку — пошаговая инструкция к вашим услугам.
Вариант 1
Берете ребенка (девочку с длинными или средними волосами).
Усаживаете на стул.
Даете в руки планшет с играми.
Вариант 2 мало чем отличается от предыдущего:
Включаем мультики.
Показываем готовый результат.
А вот и схемы плетения с фото инструкцией для мамочек, желающих сделать красивую прическу своей дочурке.
Объемный колосок на бок , начиная со лба. Плетем по схеме, которую мы уже не раз проходили, добавляя с боку прядки и скручивая их в виде жгутов.
Если раньше коса была неотъемлемым атрибутом у женщин на Руси, сегодня ее заплетают стильные девушки, в том числе голливудские звезды. Однако история колоска насчитывает гораздо больше столетий, чем кажется на первый взгляд. Еще в древнеегипетских летописях описывались способы плетения высоких причесок для знатных дам. Самые оригинальные из них держались в строжайшем секрете, который обученные рабы уносили с собой в могилу. Зачастую по косе колоском можно было узнать о количестве земель и подчиненных у богатой женщины. Длинные волосы всегда были признаком аристократического происхождения и богатства, поэтому каждая современная девушка, желающая возвысить себя в глазах окружающих, должна отрастить роскошную шевелюру и научиться плести косы колоском.
Классический колосок
Способ плетения косички колосок красиво смотрится на волосах разной длины: короткие можно в конце плетения спрятать, а длинные, наоборот, распустить. Основная схема плетения остается предельно простой: классическая косичка начинается сверху, от самой макушки.
Отделите небольшую часть волос от общего количества и разделите на три одинаковые пряди. Скрестите левую часть с правой через центральную прядь, словно вы начинаете плести обыкновенную русскую косу. Дальше выполняя аналогичные движения, добавляйте с каждой из боковых сторон по одному локону. Продолжайте плетение до тех пор, пока не дойдете до самого низа.
Схема для тупых
Коса «Рыбий хвост»
Любое плетение кос требует небольшой предварительной подготовки. Волосы необходимо тщательно расчесать массажной щеткой и увлажнить спреем или кондиционером. Эта незамысловатая процедура не позволит волосам спутываться в процессе плетения и придаст им здоровое сияние. Второе название «рыбий хвост» колосок получил за соответствующую ассоциацию с конечностью рыбы. Несмотря на то, что коса выглядит сложной и запутанной, сплести ее не составит труда.
Схема для тупых и еще тупее
Расчешите волосы назад и сделайте ровный пробор. Теперь вам по очереди необходимо брать по одной маленькой пряди с каждой из сторон, перекрещивая ее. Таким образом, каждая новая правая прядь накладывается поверх левой и крепко затягивается. В отличие от классической «русской» косы, которая основывается на переплетении трех прядей, в рыбьем хвосте участвуют только две. Рабочими остаются участки волос по бокам, не задействованные ранее в плетении косы. После того, как коса будет затянута резинкой или заколкой, ее следует растянуть в разные стороны и зафиксировать лаком, чтобы она выглядела объемнее.
Колосок наизнанку
Освоив плетение классического колоска, вам следует обратить внимание на эту прическу, словно вывернутую наоборот, то есть «наизнанку». Этот тип косы смотрится гораздо интереснее и объемнее, главное – не требует больших навыков. Просмотрев несложную схему один раз, даже самые недалекие в мире причесок девушки смогут заплести колосок наизнанку.
Схема колоска наизнанку для тупых
Этот вид косы можно начинать с самой макушки, что визуально удлинит ваши локоны. Зачерпните одну прядь волос и разделите ее на три тонкие одинаковые части. Вторую прядь наложите на первую крестообразным движением, а затем пропустите первую прядь аналогичным образом, наложив на третью. Далее по такому же принципу крестообразным движением последняя прядь ложится на вторую. Аналогичные движения повторяются до тех пор, пока вы не доплетете косу полностью. Она может не только плестись от макушки, но и украшать прическу сбоку, играя роль ободка из натуральных волос.
Колосок зигзагом
Коса необычной формы – вот что поистине оригинально выглядит на любой девушке. Особенно эффектно она смотрится на длинных густых волосах. Важно соблюдать предельную аккуратность при плетении, чтобы не оставалось залысин. Кстати, при желании можно менять форму и направление зигзагов и попробовать сделать рисунки, например, звездочки или сердечки.
Для того, чтобы сплести зигзагообразную косу, следует внимательно изучить схему классического колоска. Разделите волосы на две части и начните плести пряди справа. Плетение пряди начинается после того, когда прядь будет разделена напополам. При плетении звеньев следует прихватывать другие прядь из нижнего ряда. После того, как вы дошли до противоположного края головы, переходите на плетение с другой стороны. Важно закалывать каждый ряд спицами, чтобы звенья не распались.
Прическа «Корзиночка»
Этот вид прически, бесспорно, является самым оригинальным. Он может стать палочкой-выручалочкой вместо укладки, когда времени на поход в парикмахерскую перед мероприятием совсем нет.
На макушке сделайте круглый ровный пробор и соберите его резинкой в тонкий хвостик. Впоследствии резинка станет невидимой под красивым круговым плетением. Разделите свободную часть волос на две части и начните плетение справа. Выделяйте по одной пряди из хвостика и вплетайте их в локоны свободной части волос стандартным способом плетения классического колоска. Доплетите половинную окружность, зафиксировав снизу остаток волос. То же самое проделайте с левой стороной. В конце можно обладательницам густой шевелюры следует собрать волосы в классическую косичку, а девушки с короткими волосами могут выполнить круговое движение колоска и спрятать косичку под резинкой от хвостика при помощи невидимки.
Отзывы
Дарья Денисова, 23 года, Пермь
Выбирала себе свадебную прическу очень долго и тщательно. Торжество состоялось летом, и распускать свои длинные волосы мне однозначно не хотелось. Тем более, за целый день на ногах локоны распались бы и висели как «сосульки». Свадьба была достаточно бюджетная, поэтому я, в отличие от своих подруг, тщательно все распланировала таким образом, чтобы не переплачивать. Маникюр я сделала себе сама, макияж – работа сестры, а шикарная прическа, в основе которой лежит колосок – шедевр моей подруги. На первый взгляд сложно поверить, что это прическа парикмахера-любителя. Она не дипломированный специалист и научилась вытворять с волосами нечто волшебное сама. Правда, когда я ей показала фото того, что именно хочу видеть у себя на голове, она ответила, что никогда не плела колосок наизнанку. Мы нашли простую схему и очень быстро разобрались. Оказалось, это так легко, что этому искусству может при желании научиться каждая.
Наталья Валерьевна, 37 лет, Белгород
С процедуры плетения косы мы начинаем каждое утро моей младшей дочери, которая в этом году поступила в первый класс. Я очень люблю свою доченьку и считаю, что она должна быть самой красивой, поэтому каждый раз экспериментирую с прическами и на неделе каждый день заплетаю ее по-новому. Мастерство плетения я освоила сама, разыскивая в интернете различные схемы. Начала обучение с обыкновенного колоска, который впоследствии стал основой для плетения прической невероятной красоты.
Кто бы мог подумать, что с волосом дочки я могу выполнять красивые плетения в форме звезды или сердечка. Это кажется сложным и запутанным на первый взгляд, но если понять принцип плетения, все становится очень просто. Для того, чтобы выполнить орнамент в виде сердца, я сделала ровный пробор дочке до макушки, и далее тонким концом расчески сделала круговое движение, которое стало основой для верхней части сердечка. Способ плетения совершенно не отличается от классического колоска. В конце свободные пряди я соединяю при помощи резинки, и у меня получается ровное красивое сердечка. Радует то, что для такой прически не требуется шпилек, которые у дочки всегда выпадали.
Диана Антонова, 19 лет, Калининград
В школе я была общепризнанной «звездой» и иконой стиля. Не хочу хвастаться, но выглядела я всегда «с иголочки» даже больше не потому, что мама покупала мне дорогую одежду, а потому, что я следила за состоянием ногтей и волос. В отличие от других одноклассниц, которые приходили с модным тогда конским хвостом или нерасчесанными длиннючими волосами, я всегда заплеталась. Причем, носить хвостик или обыкновенную косичку было совсем не в моем стиле – я любила экспериментировать. Больше всего любила вплетать косичку колосок в так называемый «бублик» на голове, когда волосы собираются в пучок, а затем дульку. Когда не было физкультуры, я часто заплетала сверху боковую косичку изнаночным колоском и собирала резинкой возле уха. Колосок выполнял роль натурального ободка. Причем, я ничуть не была похожа на Юлию Тимошенко, потому что колосок был обратным, а распущенные волосы я всегда укладывала рифленым утюжком.
Видео
Травы, осоки и камыши: полевые цветы Миннесоты
Травы, осоки и камыши
Просмотреть все травы Просмотреть все осоки Просмотреть все виды камыша
В чем разница?
Травы: семейство Poaceae
- Стержни сочлененные, полые, обычно круглые в поперечном сечении, разветвленные или неразветвленные
- Оболочки обычно открываются спереди, хотя края часто перекрываются
- Листья двухрядные (по разные стороны от стебля)
- Цветки обычно идеальные (мужская и женская части)
- Каждый цветочек завернут в 2 прицветника / чешуи
- Одно семя (зерно) на цветок
Просмотреть все травы или Найти травы
Камыши: семейство Juncaceae
- Стержни круглые или сжатые в поперечном сечении, несращенные или полые, разветвленные или несоединенные
- Листья немногочисленные, в основном прикорневые, круглые или плоские в поперечном сечении
- Ножны открытые спереди, часто с ушными раковинами
- Цветки идеальные, 3 или 6 тычинок, 6 лепестков околоцветника
- Капсулы с 3 или большим количеством семян
Просмотреть все раши
Осоки: семейство Cyperaceae
- Стержни круглые или трехсторонние в поперечном сечении, неполые и не сочлененные, обычно не разветвленные
- Обшивка обычно закрывается спереди
- Листья обычно трехрядные (в 3 столбика, если смотреть сбоку от стебля)
- Цветки мужские или женские или идеальные, каждая с одной чешуей / прицветником
- Одно семя (семянка) на цветок
- Ахены линзовидные или трехсторонние, у основания могут быть щетинки или волоски
Подразделы следуют, или Просмотреть все осоки
Осоки:
Осока видов- Стебли обычно трехсторонние, листья трехрядные
- Цветы мужские или женские; колоски полностью мужские, полностью женские или смешанные
- Цветки / семянки заключены в мешок (перигиний), открываются только на кончике
- Каждый перигиний покрыт одной шкалой; обычно по спирали
- Ахены без щетины
Просмотрите только осоки Carex или выполните поиск Carex видов
Плоскости, орехи, осоки зонтичные:
Cyperus видов- Концевые скопления из нескольких колосков; Грозди часто круглые или в форме кисточки
- Колоски обычно сплющенные
- Цветки идеальные, двухрядные (соцветия по разные стороны от колоскового стебля)
- Каждый цветочек / семянка окружен единой чешуей
- Ахены без щетины
Просмотреть только Cyperus осоки
Камыши:
Scirpus , Schoenoplectus , Bulboschoenus видов- Стержни круглые или трехсторонние в поперечном сечении, губчатые или твердые
- Стеблевые листья иногда почти безлопастные
- Грозди от нескольких к большому количеству копьевидно-эллиптических колосков, расположенные по-разному
- Листовидные прицветники или прицветники, являющиеся продолжением стебля
- Цветки совершенные, расположенные по спирали на колоске
- Ахены обычно с щетиной
Просмотреть только Bulrushes
Колючки:
Eleocharis видов- Стержни круглые или сжатые в поперечном сечении, обычно губчатые
- Листья все прикорневые, по 2 на стебле; нижняя штанга обшита
- Одиночный шип на конце стебля, обычно овально-эллиптический; чешуйчатые прицветники
- Идеальные цветы, расположенные по спирали на колосе
- Ахены с заметной «шляпкой» на конце, обычно с щетинками.
Просматривать только Spikerush
Хлопковые травы:
Eriophorum видов- Стержни круглые или трехсторонние в поперечном сечении
- Стеблевые листья иногда почти безлопастные
- Концевой пучок от нескольких до многих, ланцетно-эллиптические колоски, расположенные по-разному
- Цветки совершенные, расположенные по спирали на колоске
- Ахены с длинными гладкими волосками белого или желтовато-коричневого цвета
Просматривать только хлопок
Осока другая
- Не то, что указано выше, или другие комбинации признаков
- Ахены могут иметь щетину
- Включает: Bulbostylis , Cladium , Dulichium , Fimbristylis , Lipocarpha , Rhynchospora , Scleria , Trichophourm видов
Посмотреть другие осоки
| Briza minor , дрожащая трава (Festucoideae-Poeae).Несколько из у цветков этих трех колосков тычинки выступают. Фестукоидные травы обычно иметь более одного бисексуального цветочка, и если у них есть бесплодные соцветия, т. е. те без пестиков они расположены над одним или несколькими плодовитыми соцветиями. Цветочки колоски обычно сжаты из стороны в сторону, а не спереди назад. Когда колоски созревают и распадаются, чешуйки большей частью остаются на растении. | |||
| Bro mus willdenowii , спасательная трава, Palehua-Palikea Tr., Оаху, Гавайи, 2003. | |||
| Dactylis glomerata , ежа палочка (Festucoideae-Poeae). Колоски бывают 3-6-цветковые в этом роде; у этого вида лемма имеет жесткую ость длиной до 1,5 мм. | |||
| Dactyloctenium aegyptium, колючая трава (Festucoideae-Eragrostideae) | |||
| Eleusine indica , проволочная трава (Festucoideae-Eragrostideae).В этом роде соцветия пальчатые (здесь не показаны — см. Cynodon ниже). Примечание многоцветковые, сжатые с боков колоски, расположенные в 2 череповидных ряда. | |||
| Eragrostis sp., Лавграсс (Festucoideae-Eragrostideae). Обратите внимание на сбоку сжатые колоски. | |||
| Sporobolus, indicus , западно-индийский дропсид, головня (Festucoideae-Eragrostideae).Для этого рода характерны мелкие одноцветковые колоски; Плод обычно отпадает от кожуры и палеи после созревания. | |||
| Cynodon dactylon , Бермудская трава (Festucoideae-Cynodonteae). Этот род часто имеет пальчатые соцветия с сидячими колосками в 2 ряда; колоски имеют один базальный плодородный цветочек и часто последний, стерильный или рудиментарный цветочек. На втором участке хорошо видны выступающие желтоватые тычинки и красноватые перистые рыльца. Фото. Висячие тычинки, производящие обильную, сухую, тонкостенную пыльцу и рыльца, которые большие и перистые, типичны для опыляемых ветром растений и являются наиболее распространенными режим опыления среди трав. | |||
| Chloris virgata , пальчиковый перьевой (Festucoideae-Cynodonteae). Этот род имеет пальчатые соцветия; колоски вторые в 2 ряда, остистые, несущие 1 базальный, обычно несколько вздутый плодородный цветочек и 1-3 стерильных дистальных цветочка.Обратите внимание стойкие чешуйки, характерные для фестукоидных трав, на оси слева на фото. | |||
| Brachiaria subquadripara (Panicoideae-Paniceae). Нота гладкая, безостая колоски. | |||
| Cenchrus echinatus , песчаный бор (Panicoideae-Paniceae). В этом случае соцветия окружены скоплением множества сливающихся щетинок, которые очень эффективны в способствует распространению плодов за счет спутывания их в шерсти животных или перьях птиц. | |||
| Digitaria insularis , кислая трава (Panicoideae-Paniceae). Нота шелковистая опушенные цветочки и расчленение колосков ниже чешуек. | |||
| Melinus minutiflora , патока (Panicoideae-Paniceae). Стерильная лемма каждого цветочка имеет длинную ость до 1,5 см длиной. | |||
| Oplismenus compositus , (Panicoideae-Paniceae).Этот род имеет колоски попарно; чешуя с остями до 15 мм длиной. | |||
| Panicum maximum , Морская трава (Panicoideae-Paniceae). Паникоидные травы обычно имеют небольшие колоски, обычно состоящие из одиночный тычиночный или средний цветочек у основания колоска и одиночный бисексуальный цветочек, плодородный, верхний цветочек. В период созревания весь колоск обычно отделяется от растение как единое целое, включая чешуйки.Любое сдавливание цветочков в колоске является недопустимым. обычно спереди назад, а не из стороны в сторону. | |||
| Paspalum dilatatum , трава даллиса (Panicoideae-Paniceae). Примечание добавлено тычинки и рыльца. | |||
| Paspalum fimbriatum , Panama или fimbriate paspalum, трава Колумбии (Panicoideae-Paniceae). У этого рода колоски расположены в два ряда и имеют сильная дорси-вентральная компрессия.Лемма плодородного (конечного) цветочка такова: затвердевший. Как и у большинства трав Panicoid, соцветия отделяются от оси ниже чешуйки. | |||
| Pennisetum purpureum , трава слоновая, трава ворсистая (Panicoideae-Paniceae). Этот крепкий вид может достигать 15 и более футов в высоту. Колоски бывают покрыта оберткой из многочисленных листопадных щетинок длиной до 15 мм и более. | |||
| Setaria gracilis , лисохвост желтый (Panicoideae-Paniceae). Колоски покрыты 1-несколькими стойкими шершаво-усиковидными щетинками; расчленение колоски встречаются между чешуйками и переходящими щетинками. | |||
| Coix lacryma-jobi , Слезы Иова (Panicoideae-Andropogoneae). Этот родственник кукурузы натурализован на Гавайях и иногда выращивается как декоративное растение.Перламутровые плоды-украшения иногда используются для изготовления леев. | |||
| Heteropogon contortus , пили трава, борода скрученная, клубок (Panicoideae-Andropogoneae). У этого вида каждая лемма может иметь ость до 12 см. длинный; они часто скручены вместе в верхней части соцветия. | |||
| Saccharum officinarum , сахарный тростник (Panicoideae-Andropogoneae).Некогда очень важная товарная культура на Гавайях, выращивание сахарного тростника больше не используется. экономически выгодно. | |||
| Sorghum halepense , трава Джонсона (Panicoideae). Этот конкретный триба (Andropogoneae) паникоидных трав обычно имеет парные колоски (1 стебельчатый, 1 сидячий) по оси. И паникоидные, и фестукоидные травы обычно имеют лоскут Ткань называется язычком на стыке лезвия и обшивающей части листа.В в этом случае часть листа, покрывающая оболочку, была оторвана от стебля, чтобы сделать язычок более заметен. Бахрома из волосков часто связана с язычком. | |||
| Фрагмиты sp., (Arundinoideae) | |||
| Coraderia selloana, (Arundinoideae) | |||
| Oryza sativa , рис (Oryzoideae). Этот вид — один из самых важные пищевые культуры, которые используют люди, являясь основным источником крахмала для большой сегмент населения мира. | |||
| бамбук, (Bambusoideae). Бамбук — единственные древесные представители семейство трав. Они также сочетают в себе самые примитивные персонажи, встречающиеся в семье, такие как цветочки с шестью тычинками и трехстворчатые пестики. | |||
| Bambusa ср. vulgaris , бамбук. | |||
| Giga ntochloa verticillata , бамбук (Bambusoideae).Обратите внимание Колосок сравнительно крупный, соцветия с шестью выступающими тычинками. |
Poaceae — обзор | Темы ScienceDirect
Введение
Зерновые относятся к семейству Gramineae , широко известному как травы. Большинство этих растений — многолетние; однако все товарные зерновые культуры однолетние. Считается, что доисторические люди отбирали растения, дающие большие зерна за относительно короткий период времени. С тех пор человечество манипулировало зерновыми культурами, чтобы увеличить их урожай за счет лучшей адаптации к различным экосистемам, климату и почвам.Эти непрерывные усилия по селекции растений способствовали резкому увеличению мирового населения с 1900 года (примерно с 1,7 миллиарда в 1900 году до более 7,3 миллиарда в 2014 году).
Зерновые культуры обладают множеством достоинств и преимуществ. Они дают большое количество пищи на единицу площади и зрелые зерна, которые при правильном хранении не являются скоропортящимися. Кроме того, эти зерна концентрируют калории и другие питательные вещества в относительно небольшом количестве, которое поддерживает человечество с тех пор, как оно стало малоподвижным.Среди основных групп пищевых продуктов злаки, несомненно, являются крупнейшим поставщиком калорий и белков для обычного человека.
По данным Продовольственной и сельскохозяйственной организации, зерновые на сегодняшний день являются наиболее важными продуктами питания с годовым объемом производства, превышающим 2,78 миллиарда тонн в 2013 году. Зерновые культуры являются основным продуктом питания для жителей мира и основным источником кормов для животных, которые поставляют мясо и яйца. , и молочные продукты.
Среди зерновых три контрастных зерна: рис, пшеница и кукуруза дают примерно 89% от общего объема производства.Рис и пшеница почти исключительно используются в продуктах питания человека, в то время как кукуруза широко используется в качестве сырья для животных и топливного этанола. Кукуруза является зерном с наибольшим урожаем и урожайностью, за ней следуют необработанный рис и пшеница. Соединенные Штаты собирают 35% мирового производства за счет использования высокоурожайных гибридов, включая растущее число генетически модифицированных организмов (ГМО). Пшеница по-прежнему считается иконой злаков из-за ее универсальности для производства широкого спектра закваски, что возможно благодаря ее уникальным свойствам глютена.Пшеница — это злак, который больше всего потребляет калорий. Около 45,5% пшеницы производится в Китае, Индии, США и России.
В 2014 году более 75% риса-сырца было собрано в Китае, Индии, Индонезии, Бангладеш, Вьетнаме и Мьянме. Потребление риса на душу населения в развивающихся странах Азии составляет в среднем 80 кг в год — 1 , причем люди из Мьянмы, Вьетнама, Бангладеш, Камбоджи и Индонезии являются наиболее зависимыми от этого зернового зерна с годовым потреблением 195 167, 160, 147. , и 140 кг на душу населения соответственно.
В настоящее время ячмень является четвертым зерновым зерном по объему производства с мировым производством 144,7 млн тонн в 2013 году. Рожь, ячмень и овес в основном выращиваются в Европе, России и США. Эти озимые зерновые также сажают на корм или как культуры двойного назначения. Около 65% ржи собирают в Восточной Европе из-за ее адаптации к суровым зимам и бедным почвам. В этих регионах ржаную муку обычно смешивают с пшеничной мукой для производства многих традиционных видов хлеба и других хлебобулочных изделий.Аналогичным образом, большая часть ячменя и овса выращивается в США, Канаде и Европе для производства продуктов питания и кормов. Ячмень используется для соложения, пивоварения и производства дистиллированного спирта, тогда как измельченный овес или крупа используются для производства комбинированного хлеба и других хлебобулочных изделий.
Сорго в настоящее время является пятым зерновым зерном по объему производства с глобальным производством 61,4 миллиона тонн в 2013 году. Оно непосредственно используется для производства широкого спектра традиционных продуктов питания в Африке и Индии и почти исключительно используется в качестве корма для животных в США, Мексика и другие страны Латинской Америки.Сорго в качестве корма сильно конкурирует с кукурузой, хотя считается, что его питательная ценность немного ниже.
Просо выращивают в качестве натуральных культур из-за их морозостойкости и устойчивости к засухе. Эти злаки в основном производятся в развивающихся странах Африки и Азии. Основные производители — Индия, Нигерия, Нигер и Китай. Африканские страны, расположенные к югу и западу от пустыни Сахара (Мали, Буркина-Фасо, Нигер и Нигерия), традиционно зависят от проса и сорго как основных источников питательных веществ.
Наиболее важными характеристиками злаковых растений являются стебли с узлами, на которых берут начало почки и листья, чередующиеся копьевидные листья и соцветие, которое может быть колосом или метелкой. У них несколько цветков, заключенных в чешуйки, которые после оплодотворения и созревания дают однодольные крахмалистые плоды, состоящие из околоплодника, зародыша и эндосперма. Семядоли расположены в щитке зародыша и считаются первой резервной тканью, тогда как эндосперм является самой большой частью, где белок и крахмал хранятся в качестве вторичных резервных тканей.Зерновые культуры делятся в зависимости от пути фотосинтеза на две группы: растения C 3 , которые образуют трехуглеродные соединения посредством метаболического цикла Кальвина-Бенсона, в который входят рисовая пшеница, ячмень, рожь, тритикале и овес, и растения C 4 такие как кукуруза, сорго и просо, которые образуют четырехуглеродные соединения посредством кислотного метаболического пути крассуловых. Зерновые культуры C4 обычно растут в жарком климате с высокой интенсивностью света и более эффективны с точки зрения использования CO 2 и поглощения воды и питательных веществ.
Соцветия пшеницы, ячменя, ржи и тритикале представляют собой колосья, тогда как рис, сорго, овес и все просо образуют метелки. В конкретном случае кукурузы початок классифицируется как сросшаяся метелка или центральная ось. Кукуруза, рожь, сорго и просо перекрестно опыляются, тогда как пшеница, овес, ячмень и рис — самоопыляемы.
Плод злаков ботанически называется зерновка и состоит из трех основных анатомических частей: околоплодника, эндосперма и зародыша. Некоторые виды теряют чешуйки или шелуху во время сбора урожая, а другие плотно удерживают эту защитную оболочку.Кукуруза, сорго, пшеница, рожь, тритикале и перловое просо являются примерами голых зерновок, тогда как рис, овес и ячмень — покрытые зерновки.
Околокарпий или отруби — это внешний слой, который покрывает ядро и содержит большую часть волокна, тогда как эндосперм является основной анатомической частью с точки зрения количества и использования пищи. Он может составлять до 75% от общей массы зерна, в основном состоит из крахмала и белка и практически не содержит клетчатки. Крахмал хранится в гранулах, которые классифицируются как простые или сложные.Из зерновых только рис и овес синтезируют сложные гранулы крахмала. Зародыш, который покрывает щиток и зародыш, содержит большую часть масла, а также богат белками и витаминами. Щиток или семядоли и эндосперм считаются первой и второй резервными тканями соответственно. Эти анатомические части содержат питательные вещества, необходимые для прорастания.
подавитель сидячих колосков1 Функции пути ветвления ветвей, контролирующие детерминантность меристем у кукурузы | Физиология растений
Аннотация
Колосок, представляющий собой короткую ветвь, несущую соцветия, является фундаментальной единицей архитектуры соцветий травы.У большинства трав колоски поодиночке расположены на соцветиях. Однако парные колоски характерны для Andropogoneae, трибы из 1000 видов, включая кукурузу ( Zea mays ). Подавитель сидячих колосков1 ( Sos1 ) мутант кукурузы дает одиночные, а не парные колоски в соцветии. Следовательно, ген sos1 мог участвовать в эволюции парных колосков. В этой статье мы показываем, что Sos1 является полудоминантной антиморфной мутацией. Sos1 Мутанты имеют меньше ветвей и колосков по двум причинам: (1) образуется меньше меристем пар колосков из-за дефектов размера меристемы соцветий и (2) образующиеся меристемы пары колосков образуют одну вместо двух меристем колосков. Было исследовано взаимодействие Sos1 с мутантами ramosa , которые производят больше ветвей и колосков. Результаты показывают, что Sos1 имеет эпистатическое взаимодействие с ramosa1 ( ra1 ), синергетическое взаимодействие с ra2 и аддитивное взаимодействие с ra3 .Более того, уровни мРНК ra1 снижены у мутантов Sos1 , в то время как уровни мРНК ra2 и ra3 остаются неизменными. Основываясь на этих генетических исследованиях и исследованиях экспрессии, мы предполагаем, что sos1 функционирует в ветви ra1 пути ramosa , контролируя детерминированность меристемы.
Органогенез у растений контролируется меристемами (Steeves and Sussex, 1989). Органы образуются в периферической зоне меристемы, в то время как в центральной зоне сохраняется пул клеток, которые не дифференцируются, что позволяет меристеме поддерживать себя.Меристемы определяются по их детерминированности, идентичности и положению (McSteen et al., 2000). Детерминантные меристемы продуцируют ограниченное количество структур перед окончанием, тогда как неопределенные меристемы обладают потенциалом продолжать продуцировать органы неопределенно долго (Bortiri and Hake, 2007; Sablowski, 2007). Апикальные меристемы у многих видов индетерминантные. Апикальная меристема побега (САМ) дает начало вегетативному побегу, а апикальная меристема соцветия дает начало цветущему побегу.Пазушные меристемы, образующиеся в пазухах зачатков листьев, могут быть неопределенными и давать ветви или могут быть детерминантными и давать начало цветкам.
В развитии соцветий кукурузы ( Zea mays ) существует несколько типов пазушных меристем, которые различаются по своей детерминированности, что приводит к сильно разветвленному соцветию (Irish, 1997a; Bortiri and Hake, 2007; Barazesh and McSteen, 2008b). Созревшее мужское соцветие состоит из главного колоса и нескольких длинных боковых ветвей, которые покрыты короткими ветвями, называемыми парами колосков (рис.1AРис. 1.
Sos1 фенотип кисточки и уха. A, фотография зрелых кисточек до цветения. Обычные кисточки имеют длинные ветки у основания главного шипа. Пары колосков покрывают ветви и главный колос. Мутанты Sos1 / + имеют редкий вид из-за образования меньшего количества ветвей и колосков. Sos1 / Sos1 мутанты имеют очень мало ветвей и одиночных колосков. B, количественное определение количества веток в кисточке. C: Количественное определение количества парных (серая полоса) по сравнению с одиночными (белыми) колосками на главном колоске кисточки.D. Фотография зрелых колосьев после открытого опыления. Нормальные уши имеют парные ряды ядер. Уши Sos1 / + и Sos1 / Sos1 имеют меньшее количество рядов ядер в ухе. E, Количественная оценка количества ядер на ухо. F, Количественное определение количества рядов ядер на ухо. Sos1 / Sos1 ушей меньше половины количества рядов по сравнению с обычными братьями и сестрами. Все графики являются средними ± стандартными значениями. [Цветную версию этого рисунка см. В статье в Интернете.]
Рисунок 1.
Sos1 фенотип кисточки и ушка. A, фотография зрелых кисточек до цветения. Обычные кисточки имеют длинные ветки у основания главного шипа. Пары колосков покрывают ветви и главный колос. Мутанты Sos1 / + имеют редкий вид из-за образования меньшего количества ветвей и колосков. Sos1 / Sos1 мутанты имеют очень мало ветвей и одиночных колосков. B, количественное определение количества веток в кисточке. C: Количественное определение количества парных (серая полоса) по сравнению с одиночными (белыми) колосками на главном колоске кисточки.D. Фотография зрелых колосьев после открытого опыления. Нормальные уши имеют парные ряды ядер. Уши Sos1 / + и Sos1 / Sos1 имеют меньшее количество рядов ядер в ухе. E, Количественная оценка количества ядер на ухо. F, Количественное определение количества рядов ядер на ухо. Sos1 / Sos1 ушей меньше половины количества рядов по сравнению с обычными братьями и сестрами. Все графики являются средними ± стандартными значениями. [Цветную версию рисунка см. В статье в Интернете.]
; Киссельбах, 1949).Колоск определяется как короткая ветвь с двумя листовидными чешуйками, окружающими соцветия (Clifford, 1987). Во время развития соцветия меристема верхушечного соцветия (IM) производит пазушные меристемы, называемые меристемами ветвей (BM), которые являются неопределенными и производят ветви. После того, как сделано несколько ветвей, IM резко переключается на производство меристем пар колосков (SPM), которые являются детерминированными, поскольку они производят два колоска. Меристемы колосков (SM) также детерминированы, поскольку они производят две цветочные меристемы (FM), которые затем производят цветочные органы.Хотя у кукурузы есть отдельные мужские и женские соцветия, называемые кисточкой и колосом соответственно, раннее развитие похоже, за исключением того, что колосья не дает ветвей.Были предложены две модели, чтобы объяснить, как SPM производит два SM, а SM производит два FM (Irish, 1997a, 1998; Chuck et al., 1998; Kaplinsky and Freeling, 2003). В модели латерального ветвления SPM продуцирует две SM посредством бокового ветвления, а остаточная меристема остается между двумя SM (Chuck et al., 1998). В модели преобразования SPM инициирует SM, а затем оставшийся SPM преобразуется в SM (Irish, 1997a, 1998).Сильную поддержку модели конверсии недавно предоставили эксперименты, в которых нормальные растения кукурузы обрабатывали ингибиторами транспорта ауксина, которые ингибировали инициацию SM, но не ингибировали превращение SPM в SM, что приводило к образованию одиночных колосков (Wu and McSteen , 2007).
Супрессор сидячих колосков ( Sos1 ) мутант производит одиночные, а не парные колоски в соцветии (Doebley et al., 1995). У нормальных растений один колосок прикрепляется к позвоночнику соцветия ножкой и называется колоском на ножке, а другой колоск, не имеющий цветоножки, называется сидячим колоском.У мутантов Sos1 сидячий колоск не образуется, в результате чего образуются единичные колоски на ножке. Следовательно, у мутантов Sos1 SPM продуцирует один SM вместо двух и, следовательно, более детерминирован, чем обычно.
Детерминированность SPM положительно регулируется путем ramosa (McSteen, 2006; Kellogg, 2007). ramosa1 ( ra1 ) кодирует фактор транскрипции цинкового пальца, экспрессируемый в основании SPM (Vollbrecht et al., 2005). У мутантов ra1 кисточка и початок сильно разветвлены, потому что SPM становятся неопределенными и вырастают, давая дополнительные колоски (Gernart, 1912; Vollbrecht et al., 2005). ra1 действует ниже ra2 , который кодирует фактор транскрипции LOB-домена, экспрессируемый в зачатке BM, SPM и SM (Bortiri et al., 2006a). ra1 также действует ниже ra3 , который кодирует трегалозо-6-фосфатфосфатазу (Satoh-Nagasawa et al., 2006). Интересно, что хотя ra2 — и ra3 -подобных генов присутствуют в других злаках, таких как рис ( Oryza sativa ), ra1 было идентифицировано только у Andropogoneae, клады, к которой принадлежит кукуруза.
У подавляющего большинства злаков, включая рис, ячмень ( Hordeum vulgare ) и пшеницу ( Triticum aestivum ), колоски образуются поодиночке (Clifford, 1987). Производство парных колосков характерно для Andropogoneae, племени из 1000 видов, включая кукурузу, мискантуса , сахарный тростник ( Saccharum officinarum ) и сорго ( Sorghum bicolor ; Kellogg, 2000, 2001; Grass Phylogeny Working Group, 2001).В племени Paniceae, которое является сестрой Andropogoneae, у большинства видов есть одиночные колоски, но некоторые виды производят парные колоски, а другие производят колоски, соединенные с щетинками, которые представляют собой модифицированные колоски (Kellogg, 2000; Doust and Kellogg, 2002). . Следовательно, парный колоск является производным признаком, хотя неясно, когда этот признак возник во время эволюции Panicoideae. Было высказано предположение, что ra1 могло быть задействовано для эволюции признака парных колосков (Vollbrecht et al., 2005; Макстин, 2006). Поскольку мутанты Sos1 производят одиночные, а не парные колоски, ген sos1 также может быть вовлечен в эволюцию парных колосков.
В этой статье мы используем количественный анализ, сканирующую электронную микроскопию (SEM) и гистологию, чтобы показать, что мутация Sos1 вызывает дефекты в развитии IM, BM и SPM как в кисточке, так и в ухе. Мы используем анализ дозировки, чтобы показать, что Sos1 является антиморфом, то есть доминантно-отрицательной мутацией.Более того, генетический анализ и анализ экспрессии предоставляют доказательства того, что ген sos1 действует в ветви ra1 пути ramosa , контролируя детерминированность меристемы.
РЕЗУЛЬТАТЫ
Аллель Sos1-Reference возник спонтанно (Doebley et al., 1995). Семена были получены из кукурузного генетического кооператива и подверглись 6-кратному обратному скрещиванию с получением B73. Ранее сообщалось, что мутант отображается на коротком плече хромосомы 4 (Doebley et al., 1995). Для создания картирующей популяции Sos1 (на фоне B73) скрещивали с генетическим фоном Mo17 и скрещивали с Mo17. Маркеры простого повтора последовательности (SSR) использовали для дальнейшего определения местоположения Sos1 с точностью до 4 см umc1294 в ячейке 4.02. Поскольку маркер umc1294 был полиморфен между B73 и исходным фоном, на котором возник Sos1 , этот маркер использовали для генотипирования Sos1 на фоне B73.Генотипирование в сочетании с анализом фенотипа определили, что мутация Sos1 была полудоминантной, а не доминантной, как сообщалось ранее (Рис. 1; Doebley et al., 1995).
Sos1 Мутанты производят меньше ветвей и колосковМутанты Sos1 имеют дефекты кисточки и уха. В кисточке мутантных растений Sos1 давали меньше ветвей и колосков (рис. 1, A – C). Семьи, сегрегированные по Sos1 , были генотипированы, и количество ветвей и колосков было подсчитано у нормальных братьев и сестер по сравнению с растениями, гетерозиготными и гомозиготными по Sos1 .Эти результаты показали, что мутанты Sos1 / + и Sos1 / Sos1 давали меньше ветвей, чем обычно (рис. 1B). Для анализа дефектов колосков отдельно подсчитывали количество парных колосков по сравнению с одиночными. В то время как у нормальных растений были парные колоски, у мутантов Sos1 / + и Sos1 / Sos1 было больше одиночных, чем парных колосков (рис. 1C).
Мутация Sos1 также затронула ухо. Нормальные уши не разветвлены, но дают пары колосков (Kiesselbach, 1949).Каждый колоск дает один цветочек (нижний цветочек обрывается), который при опылении дает одно ядро (Cheng et al., 1983). Колоски в колосе сидячие, поэтому спаривание не очевидно, но оно приводит к четному количеству рядов ядер в зрелом колосе. У мутантов Sos1 в ухе было меньше ядер (рис. 1D). Количественная оценка показала, что общее количество ядер было уменьшено у мутантов Sos1 / + и Sos1 / Sos1 по сравнению с нормальными ушами (рис.1E). Также наблюдалось уменьшение количества рядов ядер с мутантами Sos1 / + , дающими переменное количество рядов, и мутантами Sos1 , дающими менее половины числа рядов нормальных братьев и сестер (рис. 1F).
Чтобы определить основу развития для производства меньшего количества ветвей и колосков у мутантов Sos1 , СЭМ-анализ был выполнен на развивающихся кистях и ушах. В нормальных соцветиях ВП производятся из ИМ на верхушке соцветия (рис.2АРис. 2. СЭМ
соцветий Sos1 . От А до С, СЭМ ушей 8-недельного возраста. A, Нормальное ухо производит ряды SPM на кончике уха, которые переходят в производство парных SM ниже кончика. B, Sos1 / + ухо, производящее одиночный или парный SM. C, Sos1 / Sos1 ухо, производящее одиночный SM. От D до F, большее увеличение SM в ухе. D, Нормальное ухо с парным СМ. Наружная чешуйка (og) и внутренняя чешуя (ig) являются первыми органами, производимыми SM.E, Sos1 / + ухо, производящее одиночные или парные SM. F, Sos1 / Sos1 ухо с одиночным SM. G, обычная кисточка, создающая парные SM. H, Sos1 / + кисточка с парным SM вверху и одиночным SM у основания. I, Sos1 / Sos1 кисточкой, производящей ряды одиночных SM. Масштабная линейка = 100 мкм м от A до C и от G до I и 50 мкм м от D до F.
Рисунок 2.
СЭМ соцветий Sos1 . От А до С, СЭМ ушей 8-недельного возраста.A, Нормальное ухо производит ряды SPM на кончике уха, которые переходят в производство парных SM ниже кончика. B, Sos1 / + ухо, производящее одиночный или парный SM. C, Sos1 / Sos1 ухо, производящее одиночный SM. От D до F, большее увеличение SM в ухе. D, Нормальное ухо с парным СМ. Наружная чешуйка (og) и внутренняя чешуя (ig) являются первыми органами, производимыми SM. E, Sos1 / + ухо, производящее одиночные или парные SM. F, Sos1 / Sos1 ухо с одиночным SM.G, обычная кисточка, создающая парные SM. H, Sos1 / + кисточка с парным SM вверху и одиночным SM у основания. I, Sos1 / Sos1 кисточкой, производящей ряды одиночных SM. Масштабная линейка = 100 мкм м от A до C и от G до I и 50 мкм м от D до F.
). Затем SPM производит два SM (рис. 2, A и D). На идентичность меристемы как SM указывает образование чешуек, защитных листоподобных органов, которые являются первыми органами, продуцируемыми SM (Cheng et al., 1983). Наружная чешуйка образуется на абаксиальной стороне SM, а внутренняя чешуйка формируется внутри внешней чешуи на адаксиальной стороне SM (рис. 2D). Во время развития уха мутанты Sos1 / + и Sos1 / Sos1 продуцировали SPM как обычно, за исключением того, что оказалось меньше рядов (рис. 2, B и C). Позднее в развитии уха, когда парные SM были видны в нормальных ушах, мутантов Sos / Sos1 имели одиночные SM, а Sos1 / + мутантов имели смесь одиночных и парных SM (рис.2, Г – Ж). Аналогичным образом, в метелке SPM нормальных растений продуцировали два SM (рис. 2G), тогда как мутанты Sos / Sos1 продуцировали одиночные SM (рис. 2I), а мутанты Sos1 / + продуцировали один или два SM (рис. 2H). Следовательно, у мутантов Sos1 SPMs продуцируют один SM вместо двух. Sos1 Мутанты имеют дефекты размера IM Если бы единственным дефектом мутантов Sos1 было образование одиночных, а не парных колосков, то можно было бы ожидать, что мутанты производят ровно половину нормального количества рядов ядра.Однако мутанты Sos1 / Sos1 продуцировали менее половины нормального числа рядов ядра (Fig. 1F; Doebley et al., 1995). Более того, мутанты Sos1 / Sos1 продуцировали менее половины нормального количества колосков в кисточке (рис. 1С). Это указывает на дополнительный дефект в инициировании SPM. Для количественной оценки дефекта в инициировании SPM был использован анализ SEM для подсчета количества SPM, инициированных по окружности IM. Это показало, что нормальные уши производили 9 или 10 SPM, Sos1 / + давали в среднем 7.37 ± 0,33 SPM, в то время как Sos1 / Sos1 произвел в среднем 6,14 ± 0,34 SPM (Рис. 3AРис. 3.
Анализ IM мутантов Sos1 . A, Количественная оценка количества SPM, продуцируемых около окружность кончика Sos1 ушей. B, Измерение ширины IM. C, Измерение высоты IM. D до F, SEM IM нормального, Sos1 / + и Sos1 / Sos1 уши. Масштабная линейка = 50 мкм м.
Рисунок 3.
Анализ IM мутантов Sos1 . A, Количественная оценка количества SPM, образовавшегося по окружности кончика ушей Sos1 . B, Измерение ширины IM. C, Измерение высоты IM. От D до F, SEM IM нормальных, Sos1 / + и Sos1 / Sos1 ушей. Масштабная линейка = 50 мкм м. Для дальнейшего анализа дефектов инициации SPM соцветия заливали воском, делали срезы и окрашивали толуидиновым синим О (ТВО).Около кончика уха были видны СЗМ кольцом по окружности соцветия. На срезах, показанных на Рисунке 4
Рисунок 4.
Поперечные срезы Sos1 ушей, окрашенных ТВО. От А до С, поперечные сечения ушей около кончика уха на стадии производства СЗМ. A, Нормальное ухо показывает девять SPM, некоторые из которых только начинают формировать SM. B, Sos1 / + ухо показывает восемь SPM. C, Sos1 / Sos1 ушей, показывающих шесть SPM. От D до F, поперечные сечения позже в развитии, когда нормальные уши дали парные SM.D — Нормальное ухо с девятью парами SM. Обратите внимание, что внешняя чешуйка (g) видна на некоторых SM. E, Sos1 / + ухо, в котором два из восьми SPM производят два SM. F, Sos1 / Sos1 ухо, в котором видны шесть одиночных SM. Все SM производят чешуйки (g). Масштабная линейка = 100 мкм м. [Цветную версию этого рисунка см. В статье в Интернете.]
Рисунок 4.
Поперечные сечения ушей Sos1 , окрашенные ТВО. От А до С, поперечные сечения ушей около кончика уха на стадии производства СЗМ.A, Нормальное ухо показывает девять SPM, некоторые из которых только начинают формировать SM. B, Sos1 / + ухо показывает восемь SPM. C, Sos1 / Sos1 ушей, показывающих шесть SPM. От D до F, поперечные сечения позже в развитии, когда нормальные уши дали парные SM. D — Нормальное ухо с девятью парами SM. Обратите внимание, что внешняя чешуйка (g) видна на некоторых SM. E, Sos1 / + ухо, в котором два из восьми SPM производят два SM. F, Sos1 / Sos1 ухо, в котором видны шесть одиночных SM.Все SM производят чешуйки (g). Масштабная линейка = 100 мкм м. [Цветную версию этого рисунка см. В статье в Интернете.]
, нормальное ухо инициировало девять SPM (рис. 4A), гетерозиготное ухо инициировало восемь SPM (рис. 4B), а гомозиготное ухо инициировало шесть SPM (рис. . 4С). Позже, когда нормальные SPM инициируют два SM (рис. 4D), некоторые из SPM у мутантов Sos1 / + инициировали два SM (рис. 4E), в то время как очень немногие из SPM у мутантов Sos1 / Sos1 инициировали два SM. (Инжир.4F). Вместо этого SPMs конвертируются непосредственно в SM, что подтверждается образованием чешуек (Fig. 4F).Производство меньшего количества SPM могло произойти из-за первичного дефекта в размере IM. Таким образом, высота и ширина IM были измерены в Sos1 и нормальных ушах с использованием анализа SEM (рис. 3, D – F). Это показало, что Sos1 / Sos1 IM были на 80% шире нормальных соцветий, хотя разница была на границе статистической значимости (рис. 3B; P = 0,059). Кроме того, Sos1 / Sos1 IM в среднем составляли 71% от роста нормальных IM, и эта разница была статистически значимой (рис.3С; P = 0,014). Sos1 / + IM были промежуточными по размеру между нормальными и Sos1 / Sos1 мутантами. Следовательно, мутанты Sos1 продуцируют меньше SPM, предположительно из-за первичного дефекта в размере IM.
Таким образом, гистологический анализ и анализ SEM показывают, что мутанты Sos1 производят меньше колосков по двум причинам: (1) количество рядов SPM меньше; и (2) SPM непосредственно преобразуются в SM, не инициируя второй SM, что приводит к образованию одиночных, а не парных колосков.
Мутация
Sos1 — это антиморфАнализ фенотипа Sos1 показал, что растения, гомозиготные по мутации Sos1 , имели более тяжелый фенотип, чем растения, гетерозиготные по Sos1 , и, следовательно, мутация является полудоминантной. Были определены четыре типа доминантных мутаций (Muller, 1932). Двумя типами мутаций с усилением функции являются мутации неоморфа, которые придают новую функцию, и мутации гиперморфа, которые вызывают повышенную экспрессию гена.Два типа мутаций с потерей функции — это гипоморф и антиморф. Мутации гипоморфа также называют гапло-недостаточными или чувствительными к дозе мутациями, потому что одной копии гена дикого типа недостаточно для функционирования. У антиморфов, которые также называют доминантно-отрицательными мутациями, мутантная копия гена нарушает функцию гена дикого типа (Veitia, 2007).
Чтобы различать эти типы мутаций, используется анализ доз, чтобы варьировать дозу копии гена дикого типа в мутантном фоне (Greene and Hake, 1994).У неоморфа не будет эффекта варьирования дозы (Freeling and Hake, 1985; Poethig, 1988), в то время как у гиперморфа увеличение количества копий дикого типа приведет к более серьезному фенотипу (Kessler et al. , 2002). С другой стороны, у гипо- и антиморфов добавление копий дикого типа будет вызывать ослабление фенотипа (Poethig, 1988; Nelson et al., 2002). Гипоморфа можно отличить от антиморфа, глядя на эффект изменения числа копий гена дикого типа на фоне дикого типа.Удаление одной копии гена дикого типа вызовет видимый фенотип, если ген был гапло-недостаточным и мутация была гипоморфной, но не в том случае, если мутация была антиморфной.
Для определения фенотипического эффекта варьирования дозы sos1 дикого типа пыльцу гиперплоидов линии транслокации B-A, TB-4Sa, скрещивали с нормальными растениями или растениями, гетерозиготными по Sos1 . Скрещивание F 1 анализировали на уровень плоидности (путем подсчета абортов пыльцы; см. «Материалы и методы») и на серьезность фенотипа (путем подсчета процента одиночных колосков в кисточке и количества ядер в ухе. ).Эти результаты показали, что Sos1 растений с дополнительной копией короткого плеча хромосомы 4 ( Sos1 / + / + , гиперплоид) имели более слабый фенотип, чем растения, у которых отсутствовала нормальная копия короткого плеча хромосомы 4 ( Sos1 / — , гипоплоидный), поскольку у них был более высокий процент парных колосков (Рис. 5AРис. 5.
Анализ дозировки. A, Процент парных (серая полоса) по сравнению с одиночными (белыми) колосками в Sos1 растений, содержащих две копии хромосомы 4S дикого типа (гиперплоид) или без короткого плеча хромосомы 4 (гипоплоид).B. Процент парных (серых) по сравнению с одиночными (белыми) колосками в нормальных растениях, содержащих три копии хромосомы 4S дикого типа (гиперплоид) или одну копию хромосомы 4S дикого типа (гипоплоид).
Рисунок 5.
Анализ дозировки. A: процент парных (серая полоса) по сравнению с одиночными (белыми) колосками у растений Sos1 , содержащих две копии хромосомы 4S дикого типа (гиперплоид) или отсутствующих короткое плечо хромосомы 4 (гипоплоид). B. Процент парных (серых) по сравнению с одиночными (белыми) колосками у нормальных растений, содержащих три копии хромосомы 4S дикого типа (гиперплоид) или одну копию хромосомы 4S дикого типа (гипоплоид).
). Аналогичные результаты наблюдались и в ухе (данные не показаны). Эти результаты показывают, что Sos1 не является неоморфом, поскольку имел место эффект варьирования дозировки гена, а не гиперморф, поскольку гиперплоид не был более серьезным, чем гипоплоид.Чтобы определить, был ли Sos1 гипоморфной или антиморфной мутацией, мы создали серию доз для хромосомы 4S на генетическом фоне дикого типа. Нормальные растения, у которых отсутствует одна копия короткого плеча хромосомы 4, не имеют фенотипа Sos1 .Как в гипоплоиде (+/-), так и в гиперплоиде (+ / + / +) были в основном парные колоски (рис. 5B). Этот результат показывает, что ген sos1 дикого типа не является гапло-недостаточным и, следовательно, мутация Sos1 не является гипоморфом. Таким образом, мутация Sos1 , вероятно, является антиморфной или доминантно-отрицательной мутацией.
Sos1 Подавляет ra1 Мутантный фенотип Sos1 Мутанты имеют одиночные, а не парные колоски, потому что SPM продуцируют один SM вместо двух.Следовательно, SPM более детерминирован, чем обычно, у мутантов Sos1 . Следовательно, ген sos1 можно рассматривать как негативный регулятор детерминированности SPM. У мутантов ra1 SPM являются неопределенными, что приводит к образованию сильно разветвленных кисточек и ушей (Рис. 6, A и CРис. 6.
Генетическое взаимодействие между Sos1 и ra1 . A, Фотография зрелая кисточка всех генетических классов семейства F 2 , разделяющая на Sos1 и ra1 .B. Классификация количества различных типов подмышечных структур, создаваемых кисточкой. C. Фотография зрелых ушей всех генетических классов семейства F 2 , разделяющих на Sos1 и ra1 . D к G, СЭМ соцветия колоса. D, нормальное ухо, показывающее ряды парных SM. E, ra1 ухо, которое сильно разветвлено, так как все SPM преобразованы в ответвления. Ф, Сос1 / +; ra1 / ra1 ухо, в котором SPM производят дополнительные SM, но не так много, как ra1 .Г, Сос1 / Сос1; ra1 / ra1 колос, в котором SPM в основном дают одиночные колоски. На одной стороне наконечника нет SPM (стрелка). Масштабная линейка = 100 мкм м.
Рисунок 6.
Генетическое взаимодействие между Sos1 и ra1 . A, фотография зрелой кисточки всех генетических классов семейства F 2 , разделяющих на Sos1 и ra1 . B. Классификация количества различных типов подмышечных структур, создаваемых кисточкой.C. Фотография зрелых ушей всех генетических классов семейства F 2 , разделяющих на Sos1 и ra1 . D к G, СЭМ соцветия колоса. D, нормальное ухо, показывающее ряды парных SM. E, ra1 ухо, которое сильно разветвлено, так как все SPM преобразованы в ответвления. Ф, Сос1 / +; ra1 / ra1 ухо, в котором SPM производят дополнительные SM, но не так много, как ra1 . Г, Сос1 / Сос1; ra1 / ra1 колос, в котором SPM в основном дают одиночные колоски. На одной стороне наконечника нет SPM (стрелка).Масштабная линейка = 100 мкм м.
; Гернарт, 1912; Vollbrecht et al., 2005). Ген ra1 интерпретируется как положительный регулятор детерминированности SPM (Vollbrecht et al., 2005). Поскольку мутанты Sos1 и ra1 имеют противоположные эффекты на детерминированность SPM, мы проверили, действуют ли соответствующие гены по одному и тому же пути, создав двойные мутанты.В кисточке, Сос1 / Сос1; Двойные мутанты ra1 / ra1 были менее разветвленными, чем одиночные мутанты ra1 (рис.6А). Чтобы количественно оценить эффекты обеих мутаций, ветви на кистях были удалены, классифицированы и подсчитаны с использованием схемы, аналогичной той, которая использовалась для анализа ra1 (рис. 6B; Vollbrecht et al., 2005). Нормальные растения производят ветви, прежде чем резко переключиться на производство пар колосков. ra1 мутанты продуцируют несколько промежуточных звеньев между ветвями и парами колосков (Vollbrecht et al., 2005). Эти промежуточные продукты включают «смешанные ветви», которые представляют собой ветви со смесью одиночных и парных колосков; «Колосковые мультимеры» — ветви с одиночными колосками; и «тройные колоски».Наш анализ подтвердил, что, как и в предыдущем отчете (Vollbrecht et al., 2005), мутанты ra1 имели градацию фенотипа от ветвей до пар колосков с отсроченным переключением на идентичность SPM по сравнению с нормой (рис. 6B). . Было обнаружено, что ветви в Sos1 мутантов являются смешанными ветвями, а не истинными ветвями (рис. 6B). Sos1 мутанты давали меньше типов ветвей в целом и, следовательно, имели более ранний переход к идентичности SPM, чем обычно. В Сос1 / Сос1; ra1 / ra1 , было подавление фенотипа ra1 .Ветви были заменены смешанными ветвями, и было меньше всех типов ветвей по сравнению с одиночными мутантами ra1 (рис. 6В).
Уменьшение количества типов филиалов в Сос1 / Сос1; Двойные мутанты ra1 / ra1 потенциально могут быть объяснены тем фактом, что мутантов Sos1 продуцируют меньше SPM (Fig. 3A). Итак, мы также оценили процент типов ветвей по сравнению с общим количеством подмышечных структур, создаваемых кисточкой. В этом случае мутация Sos1 все еще оказывала подавляющее действие на фенотип ra1 , поскольку процент всех типов ветвей снижался с 41.8% в ra1 одиночных мутантов до 18,7% в Sos1 / Sos1; Двойные мутанты ra1 / ra1 . Следовательно, даже с учетом продукции меньшего количества SPM мутантами Sos1 , ветвление в двойной мутантной метелке подавлялось.
Подавление фенотипа ra1 Sos1 было даже более очевидным в ухе, чем в кисточке. Уши растений Sos1 / +; ra1 / ra1 были менее разветвленными, чем ra1 , и даже инициировали некоторые жизнеспособные ядра, что случается очень редко у одиночных мутантов ra1 .Кроме того, в Sos1 / Sos1; У двойного мутанта ra1 / ra1 ветвление было почти полностью подавлено. В самых крайних примерах уши выглядели как Sos1 , за исключением того, что они были меньше и более бесплодны (рис. 6C).
Чтобы исследовать онтогенетическую основу подавления ra1 с помощью Sos1 , SEM-анализ был выполнен на ушах семей, сегрегированных по обеим мутациям. В нормальных ушах SPM производит два SM (рис. 6D), в то время как в ra1 ушах каждый SPM становится неопределенным и разветвляется, чтобы непрерывно производить SM повторяющимся образом (рис.6E). Сос1 / +; ra1 / ra1 были подавлены по сравнению с ra1 (рис. 6, E и F). Кончик напоминал Sos1 , но у основания уха SPM разветвлялся, чтобы продуцировать множественные SM, хотя и не так много, как у мутантов ra1 (Fig. 6F). В тяжелых случаях Сос1 / Сос1; ra1 / ra1 дал уши, похожие на Sos1 (рис. 6G). Более того, у двойных мутантов иногда наблюдалось бесплодное пятно вдоль стороны уха (стрелка, рис. 6G), фенотип, который не наблюдался ни у одного из одиночных мутантов.Этот бесплодный фенотип также был виден в зрелом ухе (рис. 6С). Следовательно, мутация Sos1 подавляла фенотип мутации ra1 как в кисточке, так и в ухе.
сос1; ra2 Двойные мутанты обладают синергетическим фенотипом ra2 также работает в детерминировании SPM. У мутантов ra2 SPM становится неопределенным и дает дополнительные ветви и колоски в кисточке (Рис. 7AРис. 7.
Генетическое взаимодействие между Sos1 и ra2 . A, фотографии зрелой кисточки всех генетических классов семейства F 2 , разделяющего на Sos1 и ra2 . B. Классификация количества различных типов подмышечных структур, создаваемых кисточкой. В, Увеличенное изображение отдельных ветвей, произведенных в Сос1 / +; ra2 / ra2 кисточки двойных мутантов. D, крупный план абортированных колосков, произведенных в Sos1 / Sos1; ra2 / ra2 кисточки двойных мутантов.E, фотография зрелого уха всех генетических классов семейства F 2 , разделяющих на Sos1 и ra2 . F, вид крупным планом подмышечных структур, образовавшихся в ушах семейства двойных мутантов. Нормальный и ra2 дают два ядра в ряду, мутантов Sos1 дают одно ядро в ряду, Sos / +; ra2 / ra2 производит длинные ответвления с несколькими ядрами, Sos1 / Sos1; ra2 / ra2 дает длинные стерильные ветки без ядер. G к J, СЭМ соцветия початка.G — Нормальное ухо, показывающее ряды парных SM. H, ra2 ухо, в котором SPM образуют три SM или случайные ответвления. И, Сос1 / +; ra2 / ra2 колос, в котором СПМ образуют удлиненные ответвления. J, Сос1 / Сос1; ra2 / ra2 колос, в котором SPM образуют бесплодные удлиненные ветви. Масштабная линейка = 100 мкм м.
Рисунок 7.
Генетическое взаимодействие между Sos1 и ra2 . A, фотографии зрелой кисточки всех генетических классов семейства F 2 , разделяющего на Sos1 и ra2 .B. Классификация количества различных типов подмышечных структур, создаваемых кисточкой. В, Увеличенное изображение отдельных ветвей, произведенных в Сос1 / +; ra2 / ra2 кисточки двойных мутантов. D, крупный план абортированных колосков, произведенных в Sos1 / Sos1; ra2 / ra2 кисточки двойных мутантов. E, фотография зрелого уха всех генетических классов семейства F 2 , разделяющих на Sos1 и ra2 . F, вид крупным планом подмышечных структур, образовавшихся в ушах семейства двойных мутантов.Нормальный и ra2 дают два ядра в ряду, мутантов Sos1 дают одно ядро в ряду, Sos / +; ra2 / ra2 производит длинные ответвления с несколькими ядрами, Sos1 / Sos1; ra2 / ra2 дает длинные стерильные ветки без ядер. G к J, СЭМ соцветия початка. G — Нормальное ухо, показывающее ряды парных SM. H, ra2 ухо, в котором SPM образуют три SM или случайные ответвления. И, Сос1 / +; ra2 / ra2 колос, в котором СПМ образуют удлиненные ответвления. J, Сос1 / Сос1; ra2 / ra2 колос, в котором SPM образуют бесплодные удлиненные ветви.Масштабная линейка = 100 мкм м.
; Кемптон, 1923 г .; Никерсон и Дейл, 1955; Bortiri et al., 2006a). Однако ухо поражено лишь незначительно из-за нерегулярной гребли и случайных ветвей (рис. 7E). ra2 также играет уникальную роль в развитии соцветий, так как, в отличие от ra1 , ветви кисточки вертикальные, а ножки колосков удлиненные у мутантов ra2 (Bortiri et al., 2006a). ra2 предлагается действовать перед ra1 (Vollbrecht et al., 2005; Bortiri et al., 2006a).Чтобы проверить, действует ли sos1 по тому же пути, что и ra2 , были сконструированы двойные мутанты. Неожиданно Sos1 имел синергетическое взаимодействие с ra2 как в кисточке, так и в ухе (рис. 7). Чтобы определить влияние мутации Sos1 на фенотип ra2 в кисточке, ветви были удалены, классифицированы и количественно определены с использованием той же системы классификации, которая использовалась ранее для ra1 и ra2 (Vollbrecht et al., 2005; Bortiri et al., 2006a). ra2 мутанты, такие как ra1 , продуцировали промежуточные звенья между ветвями и колосками вдоль основного колоса, за исключением того, что фенотип был слабее, чем ra1 (рис. 7B; Vollbrecht et al., 2005; Bortiri et al., 2006a). Одиночные мутанты Sos1 и ra2 оказывали небольшое влияние на общее количество подмышечных структур, продуцируемых кисточкой (Рис. 7B; Bortiri et al., 2006a). Поразительно, но произошло значительное уменьшение (более чем в 3 раза) общего количества подмышечных структур, производимых Sos1 / Sos1; Двойные мутанты ra2 / ra2 по сравнению с любым одиночным мутантом (рис.7Б). Одиночные мутанты Sos1 и ra2 дают небольшое количество рудиментарных колосков с 1-2 чешуей без цветков (так называемые абортированные колоски). Подмышечные структуры, произведенные в Сос1 / Сос1; Двойные мутанты ra2 / ra2 почти полностью состояли из абортированных колосков (рис. 7, B и D). Мы интерпретируем эти фенотипы как синергетическое усиление дефектов ra2 .
В Сос1; ra2 с двойным мутантным ухом, усиление фенотипа ra2 также было очевидным.Уши ra2 мутантов лишь изредка дают ответвления (Vollbrecht et al., 2005; Bortiri et al., 2006a). Однако Sos1 / +; ra2 / ra2 и Sos1 / Sos1; Уши ra2 / ra2 были гораздо более разветвленными, чем уши ra2 (рис. 7E). Сос1 / +; ra2 / ra2 производил ветки с несколькими ядрами, а Sos1 / Sos1; Ветви ra2 / ra2 были длинными и бесплодными (рис. 7F).
Чтобы понять основу развития повышенного ветвления и стерильности Sos1; Уши двойного мутанта ra2 , СЭМ-анализ проводили на развивающихся ушах.В нормальных ушах SPMs продуцируют два SM (рис. 7G), тогда как у мутанта ra2 SPM продуцирует более двух SM и иногда появляются ответвления (рис. 7H). Sos1 / +; Мутанты ra2 / ra2 были более разветвленными, чем ra2 , причем каждый SPM ветвился несколько раз (фиг. 7I). Ветви были более вытянутыми, чем ra2 , и давали мало SM (рис. 7, H и I). В Сос1 / Сос1; ra2 / ra2 , фенотип был дополнительно усилен удлиненными ветвями вместо SPM и даже меньшим количеством SM (рис.7J). Следовательно, Sos1 усиливал фенотип мутанта ra2 как в кисточке, так и в ухе.
Сос1; ra3 Двойные мутанты имеют аддитивный фенотип ra3 также действует в детерминированности меристемы, хотя фенотип слабее, чем ra1 и ra2 (Satoh-Nagasawa et al., 2006). В кисточке мутанты ra3 дают несколько очень длинных ветвей, но не имеют дефектов детерминированности, наблюдаемых у мутантов ra1 и ra2 (данные не показаны).В ухе мутанты ra3 имеют неправильные ряды и ветви у основания уха (Рис. 8BРис. 8.
Генетическое взаимодействие между Sos1 и ra3 . A — D, Фотографии зрелых ушей от a Sos1; ra3 сегрегирующее семейство. От E до H, SEM-анализ ушей сегрегационного семейства Sos1; ra3 . Масштабная шкала = 100 μ м.
Рисунок 8.
Генетическое взаимодействие между Sos1 и ra3 .От A до D, фотографии зрелых ушей из Sos1; ra3 сегрегационная семья. От E до H, SEM анализ ушей от Sos1; ra3 сегрегационная семья. Масштабная линейка = 100 мкм м.
; Сато-Нагасава и др., 2006). Эти дефекты вызваны превращением SPM в ветви и SM, производящие дополнительные структуры, включая дополнительные FM (Fig. 8F; Satoh-Nagasawa et al., 2006). В Сос1 / +; Уши двойного мутанта ra3 / ra3 , ветви образовывались у основания уха, а на кончике уха образовывались одиночные колоски (рис.8С). Кроме того, аналогично ra3 одиночных мутантов, Sos1 / +; Колоски ra3 / ra3 давали дополнительные FM (рис. 8G). Сос1 / Сос1; уши двойного мутанта ra3 / ra3 имели фенотип, сходный с фенотипом Sos1 / +; ra3 / ra3 , за исключением того, что фенотип Sos1 был очевиден на большей протяженности уха (рис. 8D). Анализ SEM показал, что неопределенный SPM был произведен в основании уха (рис. 8H). Таким образом, двойные мутанты имели в основном аддитивные дефекты с одиночными рядами колосков, вызванными мутацией Sos1 на кончике, и дополнительными структурами, вызванными мутацией ra3 у основания уха.Экспрессия
ra1 , ra2 и ra3 в Sos1 Мутанты Для дальнейшего тестирования роли sos1 в пути ramosa был протестирован относительный уровень экспрессии мРНК ra1 , ra2 и ra3 на мутантах Sos1 с использованием количественной обратной транскрипции в реальном времени (RT ) -ПЦР. Результаты показали, что уровни мРНК ra1 были снижены у мутантов Sos1 / Sos1 (рис.9AРисунок 9.
Экспрессия ra1 , ra2 и ra3 в мутантах Sos1 и модель роли sos1 в детерминированности SPM. От A до C, эксперименты ОТ-ПЦР в реальном времени, показывающие относительный уровень экспрессии (A) ra1 , (B) ra2 и (C) ra3 в Sos / + и Sos / Sos1 мутанты. C. Модель взаимодействия между генами Sos1 и ramosa . ra1 , ra2 и ra3 необходимы для обеспечения детерминированности SPM. Мы предполагаем, что Sos1 является отрицательным регулятором детерминированности SPM. ra2 и ra3 действуют перед ra1 , а также имеют роли, независимые от ra1 . Ас Сос1; Двойные мутанты ra1 напоминают одиночные мутанты Sos1 , sos1 располагается после ra1 . Ас Сос1; двойные мутанты ra2 обладают улучшенным фенотипом, аналогичным ra1; Двойные мутанты ra2 , sos1 , как предполагается, положительно регулируют ra1 . Сос1; Двойные мутанты ra3 обладают аддитивным фенотипом, что указывает на то, что ra3 действует независимым путем. Модель подтверждается исследованиями экспрессии, показывающими, что ra1 снижается, но ra2 и ra3 не изменились у мутантов Sos1 .
Рисунок 9.
Экспрессия ra1 , ra2 и ra3 в мутантах Sos1 и модель роли sos1 в детерминированности SPM.От A до C, эксперименты ОТ-ПЦР в реальном времени, показывающие относительный уровень экспрессии (A) ra1 , (B) ra2 и (C) ra3 в Sos / + и Sos / Sos1 мутанты. C. Модель взаимодействия между генами Sos1 и ramosa . ra1 , ra2 и ra3 необходимы для обеспечения детерминированности SPM. Мы предполагаем, что Sos1 является отрицательным регулятором детерминированности SPM. ra2 и ra3 действуют перед ra1 , а также имеют роли, независимые от ra1 .Ас Сос1; Двойные мутанты ra1 напоминают одиночные мутанты Sos1 , sos1 располагается после ra1 . Ас Сос1; двойные мутанты ra2 обладают улучшенным фенотипом, аналогичным ra1; Двойные мутанты ra2 , sos1 , как предполагается, положительно регулируют ra1 . Сос1; Двойные мутанты ra3 обладают аддитивным фенотипом, что указывает на то, что ra3 действует независимым путем. Модель подтверждается исследованиями экспрессии, показывающими, что ra1 снижается, но ra2 и ra3 не изменились у мутантов Sos1 .
). Поскольку уровни мРНК ra1 регулируются ra2 и ra3 (Vollbrecht et al., 2005; Satoh-Nagasawa et al., 2006), мы проверили, связано ли снижение экспрессии ra1 с дефектом в ra2 или ra3 выражение. Однако уровни мРНК ra2 и ra3 не были затронуты у мутантов Sos1 / Sos1 (рис. 9, B и C). Следовательно, мутация Sos1 специфически влияла на уровни экспрессии ra1 .ОБСУЖДЕНИЕ
Самый поразительный недостаток мутантов Sos1 состоит в том, что SPM инициирует один, а не два SM. Кроме того, мутанты Sos1 имеют дефекты инициации SPM, что, как мы предполагаем, связано с общим уменьшением апикального размера IM. Sos1 мутанты также дают меньше ветвей. Однако получаемые ветви не являются нормальными, а представляют собой смешанные ветви, которые более детерминированы, чем нормальные. После того, как SM продуцируются мутантами Sos1 , они обычно дают два соцветия.Однако небольшой процент прерванных колосков (колосков без цветков) проявляется позже в развитии. О рудиментарных колосках также сообщили Doebley et al. (1995). Следовательно, мутация Sos1 влияет на детерминированность всех меристем, образующихся во время развития соцветий.
Роль
sos1 в ramosa Путь к определенности меристемra1 и ra2 мутанты сильно разветвлены, поскольку SPM являются неопределенными.Следовательно, роль генов ra1 и ra2 заключается в наложении детерминированности на SPM (Vollbrecht et al., 2005; Bortiri et al., 2006a), представленных сплошной стрелкой в модели, показанной на рисунке 9D. ra2 , как предполагается, действует выше ra1 , потому что уровни мРНК ra1 снижены у мутантов ra2 , а двойные мутанты между ra2 и слабым аллелем ra1 имеют усиленный фенотип (Vollbrecht et al. ., 2005; Bortiri et al., 2006a; Сато-Нагасава и др., 2006). Кроме того, предполагается, что ra2 будет иметь дополнительные роли, независимые от ra1 (Bortiri et al., 2006a).
У мутантов Sos1 SPM более детерминированы, чем обычно, производя один SM вместо двух. Мутация Sos1 является антиморфом, который представляет собой тип доминантной мутации потери функции (точнее, антагонист функции дикого типа). Следовательно, у мутантов Sos1 отсутствие нормальной функции гена sos1 вызывает повышение детерминированности.Одна интерпретация функции дикого типа гена sos1 заключается в противопоставлении детерминированности SPM, представленной полосой на фиг. 9D. Другой способ описания этого состоит в том, что ген sos1 придает неопределенность SPM, но поскольку SPM обычно является детерминированным, мы предполагаем, что sos1 подавляет детерминированность. Поскольку ген sos1 ингибирует детерминированность SPM, в то время как гены ramosa способствуют детерминированности SPM, были сконструированы двойные мутанты для проверки генетического взаимодействия между генами sos1 и ramosa .Удивительно, но мы обнаружили разницу во взаимодействии между sos1 и ra1 , ra2 и ra3 .
Мы предлагаем модель, показанную на рисунке 9D, для учета всех данных генетического взаимодействия и экспрессии. Сос1; Двойные мутанты ra1 напоминают одиночные мутанты Sos1 . Поскольку одиночные мутанты Sos1 и ra1 имеют противоположные фенотипы, мы интерпретируем результат двойного мутанта как означающий, что ген sos1 дикого типа функционирует ниже ra1 (рис.9D; Эйвери и Вассерман, 1992). Следовательно, ra1 может придавать детерминированность SPM путем отрицательной регуляции гена sos1 . Неожиданно Сос1; Двойные мутанты ra2 имели улучшенный фенотип по сравнению с одиночными мутантами ra2 . Sos1; Двойной мутант ra2 поразительно похож на двойной мутант между ra2 и слабым аллелем ra1 (Vollbrecht et al., 2005). Одной из гипотез, объясняющих усиление фенотипа ra2 за счет мутации Sos1 , было бы, если ген sos1 положительно регулировал ген ra1 .В поддержку этой гипотезы, уровни мРНК ra1 снижены у мутантов Sos1 . Следовательно, мы предполагаем, что sos1 действует в ветви ra1 пути ramosa , обеспечивая петлю отрицательной обратной связи для контроля детерминированности SPM (Fig. 9D).
ra3 также предположительно действует выше ra1 , хотя, поскольку ra3 кодирует трегалозо-6-фосфатфосфатазу, это взаимодействие может быть непрямым (Satoh-Nagasawa et al., 2006). ra3 мутанты имеют слабые дефекты в детерминированности SPM, указывая тем самым, что ra3 также играет роль в детерминированности SPM (Satoh-Nagasawa et al., 2006). Сос1; Двойные мутанты ra3 имели в основном аддитивные дефекты, что означает, что sos1 и ra3 действуют независимыми путями.
Мутация Sos1 оказывала более подавляющее действие на фенотип ra1 в ухе, чем кисточка. Альтернативная интерпретация фенотипа метелки Sos1; ra1 будет заключаться в том, что мутации Sos1 и ra1 обладают аддитивным эффектом, что указывает на то, что sos1 действует независимо от пути ra1 .Однако снижение экспрессии ra1 у мутантов Sos1 не поддерживает независимого взаимодействия. Более того, поскольку иногда наблюдается полное подавление фенотипа ra1 в Sos1; ra1 уха, мы поддерживаем гипотезу о том, что разница между фенотипами двойных мутантов метелки и уха обусловлена различиями в модифицирующих факторах между кисточкой и ухом. Различия в выраженности фенотипов метелки и колоса обычны для мутантов по детерминантности соцветий кукурузы (Irish, 1997a, 1997b; Laudencia-Chingcuanco and Hake, 2002; Kaplinsky and Freeling, 2003; Bortiri et al., 2006а). Одно из объяснений состоит в том, что кисточка формируется в апикальном положении, а ухо — в подмышечной, поэтому их гормональная среда, вероятно, различается. Кроме того, была продемонстрирована субфункционализация повторяющихся генов между кисточкой и ухом (Mena et al., 1996). Кроме того, степень ветвления зависит от условий окружающей среды, что указывает на то, что дополнительные модифицирующие факторы могут влиять на ветвление.
Сос1; Двойной мутант ra2 оказывал синергетическое действие на ветвление в ухе.Альтернативная интерпретация: Sos1; Фенотип двойного мутанта ra2 можно считать аддитивным, если, например, мутация ra2 вызвала неопределенность SPM, но SPM не смог инициировать сидячие SM из-за мутации Sos1 . Однако, как SPM в Sos1; Двойной мутант ra2 более неопределенен, чем одиночный мутант ra2 , мы заключаем, что взаимодействие между Sos1 и ra2 не аддитивно в ухе.Кроме того, влияние мутации Sos1 на фенотип ra2 в метелке не является аддитивным.
Таким образом, хотя можно представить себе и другие интерпретации, мы отдаем предпочтение модели, представленной на рисунке 9D, потому что она объясняет все фенотипы одиночных и двойных мутантов, а также исследования экспрессии. Доминирующие отрицательные мутации могут быть вызваны мутациями факторов транскрипции, которые, например, могут димеризовать, но не связывать ДНК, или могут связывать ДНК, но не активировать транскрипцию (Veitia, 2007).Основываясь на фенотипе мутанта Sos1 , генетических взаимодействиях с мутантами ra и снижении экспрессии ra1 у мутантов Sos1 , мы предполагаем, что sos1 может кодировать фактор транскрипции, который взаимодействует с ra1 или . ra2 .
Дополнительные роли генов
sos1 и ramosa в развитии соцветийПомимо влияния на детерминированность SPM, Sos1; ra1 и Sos; Двойные мутанты ra2 имели дополнительные дефекты развития соцветий.Это означает, что соответствующие гены играют дополнительные роли в развитии, которые ранее не были обнаружены. Сос1; Двойные мутанты ra1 давали бесплодные пятна в ухе, которые не наблюдались ни у одного из одиночных мутантов. Это синергетическое взаимодействие можно объяснить функцией обоих генов в SPM. Поскольку ra1 и ra2 не выражаются в апикальном IM, мы предполагаем, что это связано с дефектом самого SPM. Мы пришли к выводу, что, поскольку гены выполняют противоположные функции в SPM, в их отсутствие SPM иногда не запускается.Этот эффект также был замечен в кисточке, поскольку было общее уменьшение количества подмышечных структур, произведенных в Sos1; двойной мутант ra1 . В Сос1; ra2 с двойными мутантами наблюдалось даже более серьезное снижение количества подмышечных структур, образующихся в кисточке. Эти результаты показывают, что гены sos1 , ra1 и ra2 играют перекрывающиеся роли в продукции SPM как в кисточке, так и в ухе.
У одиночных мутантов Sos1 , ra1 и ra2 в кисточке образовывалось небольшое количество абортированных колосков, причем мутанты ra2 имели самый сильный эффект.В Сос1; ra1 с двойными мутантами наблюдалось несколько аддитивное увеличение количества абортированных колосков в кисточке по сравнению с любым одним мутантом. Однако в Сос1; ra2 , было синергетическое увеличение количества абортированных колосков. Фактически, почти все колоски, образовавшиеся в кисточке двойного мутанта, были абортированы. Поэтому мы предлагаем, чтобы sos1 , ra1 и ra2 также работали в SM. Однако SM все еще не определены в двойных мутантах Sos1; ra3 , аналогичных одиночным мутантам ra3 , что указывает на то, что роль sos1 не зависит от ra3 в SM.
Роль
sos1 в регулировании размера IMSos1 мутанты имеют меньший апикальный IM и меньшее количество SPM. Мы утверждаем, что дефекты в инициировании SPM происходят из-за дефектов в размере IM, поскольку есть другие примеры, когда увеличение или уменьшение размера меристемы влияло на инициирование SPM. Например, узловатых1 ( kn1 ) мутантов с потерей функции, которые также производят меньше ветвей и колосков в соцветии, уменьшили размер SAM в некоторых генетических фонах, хотя разница в размере IM не наблюдалась. продемонстрировано (Kerstetter et al., 1997; Vollbrecht et al., 2000). Противоположный фенотип наблюдается у мутантов толстых кисточек1 ( td1 ) и фасциированных ушей2 ( fea2 ), которые имеют более крупный IM и продуцируют больше SPM (Taguchi-Shiobara et al., 2001; Bommert et al. ., 2005). Интересно, что один из аллелей ra3 , ra3-fae1 , изначально был идентифицирован как имеющий фасцитированный фенотип уха (Satoh-Nagasawa et al., 2006), что указывает на то, что путь ramosa может иметь косвенное влияние на IM. . abphyl1 ( abph2 ) мутанты, которые влияют на филлотаксию листа, также имеют более крупную SAM (Jackson and Hake, 1999). abph2 кодирует регулятор цитокининового ответа, который негативно регулирует передачу сигналов цитокинина (Giulini et al., 2004). Кроме того, мутации, которые влияют на уровни цитокининов в рисе, также влияют на количество колосков (Ashikari et al., 2005; Kurakawa et al., 2007). Предполагается, что дефекты у мутантов kn1 , td1 и fea2 связаны с первичным дефектом уровней цитокининов (Barazesh and McSteen, 2008b), поскольку уровни цитокинина коррелируют с размером меристемы у eudicots (Sakakibara , 2006; Kyozuka, 2007).Имеют ли мутанты Sos1 дефекты в уровнях цитокининов или передаче сигналов, еще предстоит определить. Поскольку мутанты Sos1 и влияют на размер IM и детерминированность BM и SPM, мы предполагаем, что имеется сходный основной дефект в поддержании или детерминированности как апикальных, так и подмышечных меристем.
Играет ли ауксин роль в функции
sos1 ?Одиночные колоски на ножке также характерны для мутаций, влияющих на транспорт ауксина.Например, бесплодных соцветий1 ( Bif1 ) и bif2 мутантов, у которых нарушен транспорт ауксина, образуются одиночные колоски (McSteen and Hake, 2001; McSteen et al., 2007; Barazesh and McSteen, 2008a). бесплодный стебель1 ( ba1 ), который, как предполагается, действует как выше, так и ниже по потоку транспорта ауксина, также при мутации дает единичные колоски (Ritter et al., 2002; Gallavotti et al., 2004, 2008; Wu and McSteen, 2007; Скирпан и др., 2008). Более того, обработка нормальных соцветий кукурузы ингибиторами транспорта ауксина на более поздних этапах развития приводит к образованию единичных колосков (Wu and McSteen, 2007). Следовательно, транспорт ауксина необходим для инициирования сидячих колосков. Генетический анализ показывает Sos1; Двойные мутанты bif2 похожи на bif2 (дополнительный рисунок S1). Аналогично, ba1 эпистатичен Sos1 (дополнительный рисунок S1). Это можно интерпретировать как означающее, что sos1 действует по тому же пути, что и bif2 и ba1 , поскольку Sos1 , по-видимому, не усиливает фенотип bif2 или ba1 .Однако фенотипы bif2 и ba1 настолько серьезны, что было бы трудно увидеть улучшение. Таким образом, неясно, играет ли sos1 роль в обеспечении возможности ауксина, прямо или косвенно, инициировать сидячую SM.
В заключение, мы в настоящее время занимаемся точным картированием Sos1 для идентификации гена с помощью позиционного клонирования (Bortiri et al., 2006b). Клонирование гена sos1 позволит выяснить молекулярную основу его взаимодействия с путем ramosa .Более того, клонирование sos1 даст представление о его предполагаемой роли в эволюции парных колосков. Поскольку ra1 был идентифицирован только у Andropogoneae, было бы интересно определить, кооптировалось ли sos1 в тот же момент времени во время диверсификации трав.
МАТЕРИАЛЫ И МЕТОДЫ
Рост растений и характеристика зрелого фенотипа
Аллель Sos1-Reference был получен из Центра сотрудничества генетики кукурузы (запас 427I) и подвергнут шестикратному обратному скрещиванию с генетическим фоном B73 кукурузы ( Zea mays ).Анализ зрелого фенотипа проводили на растениях, выращиваемых в Рок-Спрингс, штат Пенсильвания, летом. Растения были генотипированы как гетерозиготы или гомозиготы Sos1 с использованием SSR-маркера umc1294 , который является ближайшим генетическим маркером, идентифицированным на данный момент (umc1294F, 5′-GCC GTC AAC GGG CTT AAA CT-3 ‘и umc1294R, 5’-GCC TCC ACG TCT CTC GTC TCT T-3 ′). Для характеристики фенотипа кисточки и колосья собирали по достижении зрелости у нормальных Sos1 / + и Sos1 / Sos1 растений из сегрегированных семейств.Количество ветвей и количество парных колосков по сравнению с одиночными на главном колосе подсчитывали на кистях после цветения. Размер выборки составил 28 нормальных, 35 Sos1 / + и 33 Sos1 / Sos1 в данных, представленных на рис. 1, B и C. Число ядер подсчитывали на открытых опыленных початках. Размер выборки был 16 нормальных, 13 Sos1 / + и 10 Sos1 / Sos1 в данных, представленных на рисунках 1, E и F.
SEM и гистология
Незрелые колосья (5–20 мм) собирали с растений, выращиваемых в течение 8 недель летом в Рок-Спрингс, штат Пенсильвания.Незрелые кисточки (3–6 мм) получали от 5-недельных растений, выращенных весной в теплице с дополнительным освещением. И кисточки, и ушки были закреплены и подготовлены для SEM, как описано (Wu and McSteen, 2007). Количество SPM по окружности апикального IM определялось путем установки верхней части уха на заглушку SEM и просмотра образцов сверху ( n = 11 нормальных, 8 Sos1 / + и 7 Sos1 / Сос1 ). Для измерения размера IM образцы колосов просматривали сбоку, и ширину IM измеряли поперек меристемы непосредственно над точкой, где были видны примордии прицветника, а высоту измеряли перпендикулярно от этой линии к верхушке соцветия ( n = 7 нормальных, 7 Sos1 / + и 6 Sos1 / Sos1 ).Кроме того, кисточки и ушки заливали воском и делали срезы для окрашивания ТВО, как описано (Wu and McSteen, 2007).
Анализ дозировки
Анализ дозировки проводили путем скрещивания известных гиперплоидов линии транслокации B-A, TB-4Sa (помеченных как sugary1 , Coop Stock 421D) как самцов с нормальными растениями или растениями, гетерозиготными по Sos1 . TB-4Sa — это реципрокная транслокация короткого плеча хромосомы 4 с сверхнормальной B-хромосомой, которая из-за очень высокой скорости нерасхождения во время второго митотического деления развития пыльцы дает пыльцу с двумя или нулевыми копиями хромосома 4S (Роман, 1947; Беккет, 1978, 1994).Плоидность потомства F 1 может быть определена по скорости аборта пыльцы (Роман, 1947; Карлсон, 1994). Аборты пыльцы оценивали с помощью карманного микроскопа по утрам подряд, когда пыльники начинали отделять свежую пыльцу (Phillips, 1994). Как наблюдалось ранее для TB4Sa, прерывание пыльцы у гипоплоидных растений составляло 50%, а у гиперплоидных растений — от 2% до 10% (Carlson, 1994; D. Auger, Университет штата Южная Дакота, личное сообщение). Кисточки и ушки гипоплоидов и гиперплоидов собирали по достижении зрелости для фенотипического анализа.Подсчитывали процент парных и одиночных колосков на 15-сантиметровом участке главного колоса кисточки после цветения. Для анализа, показанного на рисунке 4, размеры образцов составляли 13 Sos1 / + / + , 6 Sos1 / — , 8 + / + / + и 4 +/- кисточек.
Анализ двойных мутантов
Все сегрегационные семьи с двойными мутантами были созданы на генетическом фоне B73 и были высажены летом в Рок-Спрингс, штат Пенсильвания, в течение двух полевых сезонов. На каждую комбинацию двойных мутантов ежегодно высаживали не менее 360 растений.Все растения были генотипированы для Sos1 с маркером SSR umc1294 , как описано выше. Каждое семейство двойных мутантов оценивали, и анализ хи-квадрат не смог отклонить ожидаемый коэффициент сегрегации (данные не показаны).
Семьи раздельные по Sos1; bif2 и Sos1; ba1 были генотипированы по мутациям bif2 и ba1 , как описано (Skirpan et al., 2008). По мере созревания подсчитывали количество веточек кисточки и колосков на главном колосе.Для анализа, показанного на дополнительном рисунке S1B, размер выборки был 2 нормальным, 4 Sos1 / + , 11 Sos1 / Sos1 , 17 bif2 / bif2 , 13 Sos1 / +; bif2 / bif2 и 13 Сос1 / Сос1; Биф2 / Биф2 . Для анализа, показанного на дополнительном рисунке S1D, размер выборки был 2 нормальных, 5 Sos1 / + , 7 Sos1 / Sos1 , 11 ba1 / ba1 , 11 Sos1 / +; ba1 / ba1 и 8. Сос1 / Сос1; ba1 / ba1 .
Семьи раздельные Sos1; ra1 и Sos1; ra2 оценивали и анализировали фенотип кисточки и ушей в зрелом возрасте (9 недель).Ветви были удалены с кисточек и классифицированы согласно Vollbrecht et al. (2005) и Bortiri et al. (2006a). Для анализа, показанного на рисунке 6B, размер выборки был 4 нормальным, 4 Sos1 / + , 3 Sos1 / Sos1 , 9 ra1 / ra1 , 10 Sos1 / +; ra1 / ra1 и 10 Sos1 / Sos1; ra1 / ra1 . Для анализа, показанного на рисунке 7B, размер выборки был 4 нормальным, 4 Sos1 / + , 6 Sos1 / Sos1 , 6 ra2 / ra2 , 8 Sos1 / +; ra2 / ra2 и Sos1 / Sos1; ra2 / ra2 .Семьи сегрегационные по СОС1; ra3 оценивали только по фенотипу ушей, поскольку фенотип кисточки ra3 был очень слабым.
Все графики показывают среднее значение ± стандартное отклонение от среднего. Значения вероятности были определены на основе двусторонних тестов Стьюдента t , выполненных в Microsoft Excel 2003.
ОТ-ПЦР в реальном времени
Тотальную РНК экстрагировали из кисточек размером 4-5 мм из нормальных, Sos1 / + и Sos1 / Sos1 растений с помощью набора NucleoSpin RNA Plant (Macherey-Nagel GmbH & Co.). От четырех до шести образцов из каждого класса использовали в качестве биологических повторов, выполняли три технических повтора для каждого биологического повтора, и весь эксперимент повторяли три раза. Синтез кДНК, ОТ-ПЦР в реальном времени и анализ выполняли, как описано (Barazesh and McSteen, 2008a). Для обнаружения экспрессии ra1 зонд Такмана был (FAM-5′-ATC CAC AGG CTG GAC AGG GCC A-3′-BHQ), а праймеры RT-PCR были ra1F (5′-GCT GGG AGG CCA CAT GAA- 3 ‘) и ra1R (5′-GTG AAG TGT ACT GTT GGT GGA TCA G-3’).Для обнаружения экспрессии ra2 зонд Такмана был (FAM-5′-TCG TCA TTA GTA GCT CCC CAG GCG C-3′-BHQ), а праймеры RT-PCR были ra2F (5′-TGC TAC TTC ATG CGG AAC CA-3 ‘) и ra2R (5′-TTA GCC ACG GAA GCG TAA GG-3’). Для обнаружения экспрессии ra3 зонд Такмана был (FAM-5′-CCG TCC ACT TCC GCT GCG TCC-3′-BHQ), а праймеры RT-PCR были ra3F (5′-CCA GGG TGG AGC ACA ACA A- 3 ‘) и ra3R (5’-CTC GTT CAC GGC ATT CCA T). Внутренний контроль для амплификации — убиквитин ( убк ).Для обнаружения экспрессии ubq зонд Taqman представлял собой (5′-FAM-AAA TCC ACC CGT CGG CAC CTC C-3 ‘BHQ), а праймеры RT-PCR были ubqF (5’-CTC TTT CCC CAA CCT CGT GTT- 3 ‘) и ubqR (5′-ACG AGC GGC GTA CCT TGA-3’).
Дополнительные данные
В онлайн-версии статьи доступны следующие материалы.
БЛАГОДАРНОСТИ
Мы благодарим Эрика Фоллбрехта за указание на сходство между Sos1; ra2 и ra1; двойные мутанты ra2 , Дэвид Барнс и Сара Хейк за первое открытие Sos1; ra2 и двух анонимных рецензентов за полезные предложения.Мы благодарим Деба Гроув из Центра нуклеиновых кислот Института Хака за проведение экспериментов ОТ-ПЦР в реальном времени, Мисси Хейзен из Центра электронной микроскопии Института Хака за обучение работе с SEM, а также Джеффри Бутербо, Джейсона Хоара, Криса Кука, Кима Филлипса, Мэтту Дэвису и Крису Хадсону за помощь в полевых условиях и за генотипирование Sos1 . Мы благодарим Maize Coop за семена Sos1 , Фрэнка Бейкера за семена гиперплоидов TB4Sa и Дона Аугера за советы по анализу дозировки туберкулеза. Благодарим Тони Омейса и Скотта Харккома за уход за растениями.Мы благодарим сотрудников лабораторий Макстина и Брауна за обсуждение и комментарии к рукописи.
ЦИТИРОВАННАЯ ЛИТЕРАТУРА
Асикари М., Сакакибара Х., Лин С.Ю., Ямамото Т., Такаши Т., Нишимура А., Анхелес Е.Р., Циан К., Китано Х., Мацуока М. (
2005
) Цитокининоксидаза регулирует производство зерна риса.Наука
309
:741
—745
Эйвери Л., Вассерман С. (
1992
) Упорядочивающая функция гена: интерпретация эпистаза в регуляторных иерархиях.Trends Genet
8
:312
—316
Баразеш С., Макстин П. (
2008
a) бесплодное соцветие1 участвует в органогенезе во время вегетативного развития и развития соцветий кукурузы.Генетика
179
:389
—401
Баразеш С., Макстин П. (
2008
b) Гормональный контроль развития соцветий травы.Trends Plant Sci
13
:656
—662
Beckett JB (
1978
) Транслокации B-A у кукурузы.I. Использование для определения местоположения генов по хромосомным плечам.J Hered
69
:27
—36
Beckett JB (
1994
) Определение местоположения рецессивных генов в плече хромосомы с транслокациями B-A. В М. Фрилинг, В. Уолбот, ред., Справочник по кукурузе. Springer-Verlag, New York, pp 315–327Bommert P, Lunde C, Nardmann J, Vollbrecht E, Running M, Jackson D, Hake S, Werr W (
2005
) толстая кисточка dwarf1 кодирует предполагаемый ортолог кукурузы из семейства рецепторов, богатых лейцином CLAVATA1 арабидопсиса. киназа.Девелопмент
132
:1235
—1245
Bortiri E, Chuck G, Vollbrecht E, Rocheford T, Martienssen R, Hake S (
2006
a) ramosa2 кодирует белок домена LATERAL ORGAN BOUNDARY, который определяет судьбу стволовых клеток в меристемах ветвей кукурузы.Растительная ячейка
18
:574
—585
Бортири Э., Хак С. (
2007
) Цветение и определенность кукурузы.J Exp Bot
58
:909
—916
Бортири Э., Джексон Д., Хейк С. (
2006
b) Достижения в геномике кукурузы: появление позиционного клонирования.Curr Opin Plant Biol
9
:164
—171
Carlson WR (
1994
) Манипуляция транслокацией B-A. В М. Фрилинг, В. Уолбот, ред., Справочник по кукурузе. Springer-Verlag, New York, pp 308–314Cheng PC, Greyson RI, Walden DB (
1983
) Инициирование органа и развитие однополых цветков в кисточке и колосе Zea mays .Am J Bot
70
:450
—462
Chuck G, Meeley RB, Hake S (
1998
) Контроль судьбы меристемы колосков кукурузы с помощью APETALA2 -подобного гена неопределенного колоска 1 .Genes Dev
12
:1145
—1154
Clifford HT (
1987
) Морфология колосков и цветков. В Т. Р. Содерстром, К. В. Хилу, К. С. Кэмпбелл, М. Е. Баркворт, редакторы, Grass Systematics and Evolution.Smithsonian Institution Press, Вашингтон, округ Колумбия, стр. 21–30Doebley J, Stec A, Kent B (
1995
) Подавитель сидячих колосков1 ( Sos1 ): доминантный мутант, влияющий на развитие соцветий кукурузы.Am J Bot
82
:571
—577
Доуст А.Н., Келлогг Е.А. (
2002
) Диверсификация соцветий в кладе паникоидных «щетинистых трав» (Paniceae, Poaceae): данные молекулярной филогении и морфологии развития.Am J Bot
89
:1203
—1222
Freeling M, Hake S (
1985
) Генетика развития мутантов, которые определяют узловатые листья кукурузы.Генетика
11
:617
—634
Gallavotti A, Yang Y, Schmidt RJ, Jackson D (
2008
) Взаимосвязь между транспортом ауксина и ветвлением кукурузы.Plant Physiol
147
:1913
—1923
Галлавотти А., Чжао К., Киозука Дж., Мили Р. Б., Риттер М., Добли Дж. Ф., Пе М. Е., Шмидт Р. Дж. (
2004
) Роль бесплодного стебля 1 в архитектуре кукурузы.Природа
432
:630
—635
Gernart W (
1912
) Новый подвид Zea mays L.Am Nat
46
:616
—622
Джулини А., Ван Дж., Джексон Д. (
2004
) Контроль филлотаксии с помощью цитокинин-индуцируемого гомолога регулятора ответа ABPHYL1.Природа
430
:1031
—1034
Рабочая группа по филогении трав (
2001
) Филогения и подсемейная классификация трав ( Poaceae ).Энн Мисс Бот Гард
88
:373
—457
Greene B, Hake S (
1994
) Использование сегментарных анеуплоидов для анализа мутантов. В М. Фрилинг, В. Уолбот, ред., Справочник по кукурузе. Springer-Verlag, New York, pp 270–273Irish EE (
1997
a) Класс II семян метелки мутации свидетельствуют о множестве типов меристем соцветий у кукурузы (Poaceae).Am J Bot
84
:1502
—1515
Irish EE (
1997
b) Экспериментальный анализ развития метелки у мутанта кукурузы Tassel seed 6 .Plant Physiol
114
:817
—825
Irish EE (
1998
) Колоски травы: острая проблема.Bioessays
20
:789
—793
Джексон Д., Хейк С. (
1999
) Контроль филлотаксии кукурузы с помощью гена abphyl1 .Девелопмент
126
:315
—323
Каплинский Н.Дж., Фрилинг М. (
2003
) Комбинаторный контроль идентичности меристемы в соцветиях кукурузы.Девелопмент
130
:1149
—1158
Kellogg EA (
2000
) Молекулярная и морфологическая эволюция Andropogoneae. В SWL Jacobs, JE Everett, eds, Grasses: Systematics and Evolution. CSIRO, Коллингвуд, Австралия, стр. 149–158Kellogg EA (
2001
) История эволюции трав.Plant Physiol
125
:1198
—1205
Kellogg EA (
2007
) Цветочные проявления: генетический контроль соцветий травы.Curr Opin Plant Biol
10
:26
—31
Kempton JH (
1923
) Наследственные признаки кукурузы XIV: разветвленные колосья.J Hered
14
:243
—252
Керстеттер Р.А., Лауденсия-Чингкуанко Д., Смит Л.Г., Хейк С. (
1997
) Мутации с потерей функции в гене гомеобокса кукурузы, узловатый1 , нарушают поддержание меристемы побегов.Девелопмент
124
:3045
—3054
Kessler S, Seiki S, Sinha N (
2002
) Xcl1 вызывает отложенные косые периклинальные деления клеток в развивающихся листьях кукурузы, что приводит к дифференцировке клеток по клонам, а не по положению.Девелопмент
129
:1859
—1869
Киссельбах Т.А. (
1949
) Структура и воспроизводство кукурузы. Лабораторная пресса Колд-Спринг-Харбор, Колд-Спринг-Харбор, Нью-Йорк, стр. 37–63Куракава Т., Уэда Н., Маэкава М., Кобаяси К., Кодзима М., Нагато Ю., Сакакибара Х., Киодзука Дж. (
2007
) Прямой контроль активности меристемы побегов с помощью активирующего цитокинин фермента.Природа
445
:652
—655
Kyozuka J (
2007
) Контроль функции меристемы побега и корня цитокинином.Curr Opin Plant Biol
10
:442
—446
Laudencia-Chingcuanco D, Hake S (
2002
) Неопределенный ген вершины цветка регулирует детерминированность и идентичность меристемы в соцветии кукурузы.Девелопмент
129
:2629
—2638
McSteen P (
2006
) Ответвление: путь ramosa и эволюция морфологии соцветий травы.Растительная ячейка
18
:518
—522
McSteen P, Hake S (
2001
) бесплодное соцветие2 регулирует развитие пазушных меристем в соцветиях кукурузы.Девелопмент
128
:2881
—2891
Макстин П., Лауденсия-Чингкуанко Д., Коласанти Дж. (
2000
) Цветочек под любым другим названием: контроль идентичности меристемы у кукурузы.Trends Plant Sci
5
:61
—66
McSteen P, Malcomber S, Skirpan A, Lunde C, Wu X, Kellogg E, Hake S (
2007
) бесплодное соцветие2 кодирует коортолог серин / треонинкиназы PINOID и требуется для органогенеза во время соцветие и вегетативное развитие кукурузы.Plant Physiol
144
:1000
—1011
Мена М., Амброуз Б.А., Мили Р.Б., Бриггс С.П., Янофски М.Ф., Шмидт Р.Дж. (
1996
) Диверсификация активности С-функции в развитии цветков кукурузы.Наука
274
:1537
—1540
Muller HJ (
1932
) Дальнейшие исследования природы и причин генных мутаций. В Д. Ф. Джонс, В. И. Менаша, ред., Труды Шестого Международного конгресса генетиков.Бруклинский ботанический сад, Бруклин, Нью-Йорк, стр. 213–255Nelson JM, Lane B, Freeling M (
2002
) Экспрессия мутантного гена кукурузы в вентральном эпидермисе листа достаточна, чтобы сигнализировать о переключении дорсовентральной оси листа.Девелопмент
129
:4581
—4589
Nickerson NH, Dale EE (
1955
) Модификации кисточки в Zea mays .Энн Мисс Бот Гард
42
:195
—211
Филлипс Р.Л. (
1994
) Классификация абортов пыльцы в полевых условиях. В М. Фрилинг, В. Уолбот, ред., Справочник по кукурузе. Springer-Verlag, New York, pp 297–298Poethig RS (
1988
) Гетерохронные мутации, влияющие на развитие побегов кукурузы.Генетика
119
:959
—973
Ritter MK, Padilla CM, Schmidt RJ (
2002
) Мутант кукурузы barren stalk1 имеет дефект в развитии подмышечной меристемы.Am J Bot
89
:203
—210
Роман Х. (
1947
) Митотическое нерасхождение в случае обменов с участием хромосомы В-типа кукурузы.Генетика
32
:391
—409
Сабловски Р. (
2007
) Цветение и определенность у Arabidopsis.J Exp Bot
58
:899
—907
Сакакибара Х. (
2006
) Цитокинины: активность, биосинтез и транслокация.Annu Rev Plant Biol
57
:431
—449
Сато-Нагасава Н., Нагасава Н., Малкомбер С., Сакаи Х., Джексон Д. (
2006
) Фермент метаболизма трегалозы контролирует структуру соцветий кукурузы.Природа
441
:227
—230
Skirpan A, Wu X, McSteen P (
2008
) Генетическое и физическое взаимодействие предполагает, что бесплодный стебель1 является целью бесплодных соцветий2 в развитии соцветий кукурузы.Завод J
55
:81
—91
Стивс Т., Сассекс I (
1989
) Модели в развитии растений. Издательство Кембриджского университета, Кембридж, ВеликобританияTaguchi-Shiobara F, Yuan Z, Hake S, Jackson D (
2001
) Ген фасцииированного уха 2 кодирует богатый лейцином рецептор-подобный белок, который регулирует пролиферацию меристемы побегов кукурузы.Genes Dev
15
:2755
—2766
Veitia RA (
2007
) Изучение молекулярной этиологии доминантно-отрицательных мутаций.Plant Cell
19
:3843
—3851
Vollbrecht E, Reiser L, Hake S (
2000
) Размер меристемы побега зависит от инбредного фона и наличия гена гомеобокса кукурузы, узловатый1 .Девелопмент
127
:3161
—3172
Фоллбрехт Э., Спрингер П.С., Гох Л., Баклер ИВЕС, Мартиенсен Р. (
2005
) Архитектура систем цветочных ветвей кукурузы и родственных трав.Природа
436
:1119
—1126
Wu X, McSteen P (
2007
) Роль транспорта ауксина во время развития соцветий кукурузы, Zea mays (Poaceae).Am J Bot
11
:1745
—1755
Заметки автора
© 2009 Американское общество биологов растений
© Автор (ы) 2009. Опубликовано Oxford University Press от имени Американского общества биологов растений.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/), которая разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии, что оригинальная работа правильно процитировано.
Структура травы
Структура травыБольшая часть этого раздела заимствована из материала Grasses или основана на нем. Нового Южного Уэльса (Wheeler et al., 1982)
Растительные символы
Вегетативная или нецветущая часть травянистое растение очень изменчиво.Растения могут вырасти до пучков или хаотично альпинисты. В то время как вегетативные признаки часто легко различить, у трав они часто чрезвычайно изменчивы, на них легко влияют климатические и пищевые условия.
Привычка
Habit описывает основной вид растения, который может быть довольно разнообразным. в траве. Это варьируется от символов основного размера до других, описывающих шероховатость или опушенность различных листовых частей растения. Цветущие стебли включены с вегетативными символами, чтобы избежать путаницы, поэтому высота стебля означает максимальную высоту растения вне зависимости от того, цветет оно или нет.
ПродолжительностьПродолжительность жизни трав непостоянна. Некоторые могут произойти только на короткое время после дождя и их циклы не могут быть привязаны к временам года (эфемерные). Другие завершают свой жизненный цикл в течение года и начинают с посевного материала каждый год (ежегодно). Третьи сохраняются в течение двух (двухгодичных) или более (многолетних) лет. Это может быть персонажем, которого сложно определить. Если растение сложно подтянуть и имеет массу корней и остатки старых побегов вокруг основания, что может хорошо быть многолетним.Если растение относительно тонкое и с тонкими корнями, оно скорее всего, будет ежегодным.
Корневища и столоныМногие травы растут в виде компактных пучков или кисточек, но другие дают побеги. которые растут горизонтально и укореняются в каждом узле. Это полезно для растения, потому что если побег сломается, он продолжит расти как отдельное растение. Если бегуны встречаются над землей, они называются столонов, и представляют собой корневищ . если они возникают под землей.Узлы столонов дают зеленые листья, в то время как маленькие коричневые чешуевидные листья образуются на узлах корневищ.
Происхождение ВсходовПобеги образуются из пазушных почек, лежащих в углу, где стебель и лист-ножны встречаются. Если новый побег вырастает между стеблем и влагалищем листа он обозначен как интравагинальный . Если он прорвет основание оболочки листа его называют экстравагинальным побегом .
КульмсСтебель — это вертикальный стебель в середине побега (или культиватора).Они твердые в узлах и часто полые по всей области междоузлий. В некоторых травах междоузлия стебля очень короткие, пока стебель не удлиняется для цветения. Стебли бывают от неразветвленных (простых) до многоветвистых.
ЛистьяТравяной лист представляет собой удлиненную структуру, возникающую в узле и состоящую из базальной цилиндрической оболочки , которая окружает стебель или молодые листья. Над ним — приплюснутый клинок с параллельным жилкованием.
Гильзы полые цилиндры разделить на одну сторону с перекрытием полей. Выступы в виде ушей (ушные раковины) часто образуются в верхней части влагалища листа. Ушные раковины могут быть редуцируется до волосистого края основания листовой пластинки или может встречаться на одной стороне только лист. Ушные раковины, хотя и являются отличительной чертой многих видов, но очень нежные. структуры и могут увядать с возрастом. Молодые листья всегда следует осматривать, когда работа с ушными раковинами.
Полоса механической ткани, часто бледная, на стыке лезвия и ножен, абаксиальная часть которого называется воротником .(Абаксиальный относится к поверхность, обращенная в сторону от оси растения). Абсциссионный слой может также развиваются, позволяя сбивать лезвие.
A язычок , в форме языка или лоскутная прозрачная мембрана или волоски, прикрепляется к поверхности листа облицовкой шток (адаксиальная поверхность) в месте соединения ножа и оболочки. Тип язычка особенно полезный идентификационный символ, поскольку он относительно постоянен внутри рода. У некоторых трав может отсутствовать язычок, например Эхинохлоа crus-galli (Скотный двор).Многие травы имеют полупрозрачные перепончатые язычки, тогда как у других язычки окаймлены волосками или имеют форму волоскового ободка.
В основании листового влагалища и пластинки вставочных меристем что для всех, кроме старых листьев, позволяет листу удлиниться после удаления лезвие из-за скашивания или выпаса скота или повреждения ветром и дождем. Интеркалярные меристемы это области, в которых клетки сохраняют свою способность делиться и производить новые клетки. Поскольку листья травы созревают от кончика лезвия до основания, меристема влагалища остается активной дольше, чем меристема лопасти.Интеркалярные меристемы также встречаются в травах у основания междоузлий стебля, где активность позволяет стебель после полегания вырастает вертикально.
Хотя форма, текстура, складки, ворсистость и т. Д. Листовой пластинки могут быть полезными диагностическими функциями, они часто изменяются в пределах одного вида или даже на одном и том же заводе. У некоторых листовых пластинок средняя жилка видна, и часто бледного цвета, у других нервы столь же заметны или незаметны. Вернация часто определяется как расположение молодого листа в бутон, здесь мы определили вернацию как расположение листа при просмотре в поперечном сечении (поперек листа).Листовые пластинки часто плоские, но могут скатываться или складываться (перекатывание или складывание — это реакция на водный стресс в некоторых футляры), а некоторые из них настолько узкие, что напоминают щетину.
Персонажи соцветий
Основные признаки соцветияСтебель травяного культиватора в репродуктивной стадии оканчивается одним или несколькими соцветий или цветущих головок. Соцветие включает в себя все структуры над последним листом стебля или основания. В некоторых тропических травах то, что сначала кажется соцветием, представляет собой сложный набор ветвей с листовой структурой, шпате или шпате , в основании каждого соцветия в отдельности.Ветви над спатеолой могут уплотняться. или модифицированы и часто бывают парами. Пятна могут быть цветными или зелеными и могут или не могут быть разделены на лезвие и ножны.
Каждое соцветие состоит из нескольких колосков , которые содержат очень мелкие цветки, расположенные на ветвях или главная ось. Колоски, составляющие соцветие травы, бывают либо прикреплены непосредственно к ветвям, в этом случае они сидячие , или прикрепляются при помощи стеблей или цветоножек.Стебель сразу под соцветие — цветонос, , продолжающийся главной осью или rachis после первой ветки или колоска. «Рашис» — это общий термин, часто используется для любой оси или ветви, к которой прикреплены колоски.
В этом ключе мы разделили соцветия по их основной форме, если смотреть.
на расстоянии. Соцветие в форме ручки или эскимо.
при осмотре на расстоянии вытянутой руки обозначается сплошным , а соцветие с
явными ветвями является сложное соцветие .Соцветия с двумя
или несколько ветвей, происходящих из одной и той же точки, имеют цифр , или если
ветви возникают почти из той же точки subdigitate .
Хотя соцветие может состоять из колосков, расположенных непосредственно на рахисе, основной вид соцветия травы — сложная метелька , обычно называется просто метелкой, со вторичным ветвлением и часто более высоким порядком ветвления. Ветвь высшего порядка может состоять из шипа с прикрепленными колосками. непосредственно, в то время как в кисть колоски прикреплены к рахису по цветоножкам.Ветви как с сидячими, так и с цветоножками технически считаются баранов , но здесь рассматриваются как кисти. Много соцветий, которые появляются на расстоянии, чтобы быть одиночным позвоночником, на самом деле являются промежуточными по типу и могут можно рассматривать как метелки с пониженным ветвлением. Если ветки короткие и скученная, метелка узкая и описывается как шиповидная , сжатый или плотный. Уменьшение количества ветвей может вызвать первичная ось с кистями или шипами.
КолоскКаждый колосков состоит из нескольких цветков или цветков, расположенных попеременно по обе стороны от центральной оси, rhachilla , над двумя пустые прицветники, чешуек . «Прицветник» — это общий термин для обозначения сильно уменьшенного лист, особенно мелкие листья, переходящие в цветочки. Приложен цветок травы двумя прицветниками, внешний на стороне от оси, названной леммой и внутренний прицветник, названный palea .Прицветники вместе с цветком, составить цветочек.
Колоски в соцветии обычно похожи по внешнему виду, но двух типов может находиться в одном соцветии или в разных соцветиях. В Ламарки стерильные колоски, не дающие плодов, смешивают с плодородными колосками.
Колоски многих родов трибы Andropogoneae рождаются в разнородных парах, один из которых плодороден (и часто сидячие) и другие ножки, стерильные и меньшие по размеру, их называют компаньонами. колоски стерильные .
Мужские и женские колоски могут находиться в отдельных соцветиях, которые часто непохожий на вид. В однодомном растении, таком как Zea mays (кукуруза), оба виды соцветий встречаются на одном и том же растении. В Zea mays самец соцветие терминальное, а самка (початок) находится в пазухах нижние листья.
У двудомного растения, такого как Spinifex sericeus, мужских и женских соцветий. находятся на разных заводах. В Cleistochloa колоски с цветами, которые нормально открытые для опыления (хазмогамные) встречаются в соцветиях одного типа, в то время как клейстогамные колоски (которые не раскрываются) встречаются в соцветиях, заключенных в в листовых ножнах.Колоски или соцветия, встречающиеся в двух формах: часто называют диморфным.
Многие травы, кроме упомянутых типов андропогоноидов, имеют колоски. расположены в группах, например Hordeum (Ячмень).
Степень зрелости колосков может варьироваться в пределах большого соцветия. Часто более старые колоски находятся ближе к вершине или середине соцветия. Желательно и обычно можно исследовать колоски на разных стадиях. зрелости при определении травы.
InvolucreКолоски или группы колосков могут быть покрыты различными структурами, которые функция в рассредоточении семян. Эти структуры включают волосы, щетину, шипы, заусенцы или стерильные ветви, и называются оберткой.
РахиллаЭта малая ось попеременно изгибается в противоположных направлениях (изгиб или зигзаг). и несет цветки в два ряда.
КожиНа каждом колоске по две чешуи на база.Чешуйки никогда не переходят в цветок или палею. Однако они могут быть изменены и различаются по размеру, форме, текстуре и т. д., и один (или редко оба) могут отсутствовать. Похоже на полосы на чешуях — это нити сосудистой ткани (вены), называемые нервов . Количество и выступание нервов варьируется между видами, но достаточно постоянный внутри вида. Глюмы могут напоминать приведенные выше леммы их и важно, рассматривая соцветия травы, уметь распознают чешуйки и таким образом идентифицируют колоски как единицы.
ЦветочкиСамая внешняя структура цветочка — это лемма , которая обычно включает другие части. При боковом сжатии лемма обычно имеет один выступ или киль , идущий по середине спины. Обычно видно, и часто постоянное количество для определенного рода или вида, несколько жилок или Нервы . Леммы часто зеленые и перепончатые, особенно у незрелых, и по мере созревания становятся более твердыми и часто менее зелеными.Есть значительный изменение размера, формы, цвета, текстуры, сжатия, волосатости и присутствия придатков, таких как awns (см. варианты). Такой варианты особенно полезны при идентификации.
У основания многих цветков (как в Aristida и Austrostipa ) представляет собой затвердевшую структуру, часто резко заостренную, которая развивается по мере созревания и хорошо видно, когда цветочек отделяется от рахиллы. Это каллуса может быть образован как из части рахиллы, так и из части леммы и полезен при распространении семян, позволяя лемме прилипать к шкурам животных, и проникнуть в почву после того, как агрегат будет рассредоточен.Острая мозоль Hordeum murinum (трава ячменя) вызывает слепоту у ягнят и ягнят травы, такие как Aristida spp. (Проволочная трава) значительный ущерб животному прячется. Твердое, обычно заостренное основание колоска (как у Heteropogon , Andropogon и родственные роды) чуть выше точки расчленения также называется мозолью. В данном случае это часть рахиллы. В Eriochloa каллус — утолщенный узел и остаток первой чешуи; в Chrysopogon входит в состав цветоножки.
Paleas обычно двухкилевые (и с двумя нервами), более мягкие и менее изменчивые. по структуре, чем леммы. У цветочка плоская сторона обращена к рахилле. в то время как кили касаются сторон леммы, таким образом охватывая цветок. Палеас бамбука и Oryza sativa (Рис) и их родственники лишь частично вложены в их леммы.
ЦветыНет чашелистиков или лепестков, но есть два или три маленькие мясистые структуры, лодукулов, , у основания яичника, считаются редуцированным околоцветником.Лодикулы — это нежные структуры. лежит между яичником и леммой (и палеа, если есть третья). В качестве цветок раскрывается (цветение) они раздуваются, раздвигая чешую и палею, чтобы позволяйте сначала пыльникам (обычно), а затем клеймам (проект за прицветниками), позволяя опыление. Protandry — состояние в у которых тычинки отрываются первыми и протогиния при появлении рыльц перед пыльниками. Когда рыльца остаются открытыми в течение нескольких часов, узелки сморщиваются, позволяя lemma и palea сомкнуться над развивающимся зерном.
Прикрепляется к основанию цветка кольцом или завитком внутри лодикул, 1-6 тычинок. У большинства трав есть 3 тычинки с длинными тонкими волокнами. Пыльники, открывающиеся через прорези, универсальны; это нити прикреплены в одной точке над основанием. Это позволяет пыльнику качаться с ветер, позволяющий пыльце разлетаться во всех направлениях.
Обычно удлиненный верхний яичник содержит один локулюс или камера, с одной яйцеклеткой , прикрепленной к основанию на бледной стороне.На верхушке завязи расположены два боковых стика (у некоторых злаков сросшиеся чтобы дать единый центральный стиль) с волосатыми рыльцами. В тучных травах рыльца бледные, у паникоидных трав обычно темные (от розового до черного).
Зерно Семяпочка травяного цветка развивается, после
оплодотворение, в семя с большим количеством крахмалистого эндосперма и зародыш, лежащий на
одна сторона. Стенка яичника, образующая околоплодник плода, в норме
слился с семенной оболочкой, и плод получил название зерновки или зерна . Sporobolus и Eleusine продуцируют семянки, у которых теста (семя
шерсть) свободна от околоплодника. Некоторые виды бамбука дают ягоды. Большинство зерен травы
заключены в лемму и палеу. Форма, поверхность, текстура, относительный
размер и положение эмбриона — полезные диагностические признаки.
Многие систематики трав считают основным («типичный») рисунок колосков, чтобы быть похожим на многоцветковые колоски травы, такие как Avena sativa (овес).Считается, что это было изменено у других видов на добавлено таких структур, как ости, волоски и т. д. и на уменьшение на количества и / или размеров частей колосков.
ДополненияВолосы, щетина, шипы и т. Д. Могут быть прикреплены к частям колосков. В некоторых Случаи таких структур окружают отдельные колоски или группы колосков. Самый очевидный придаток — ость.
Ось представляет собой длинный тонкий отросток, прикрепленный к кончику, спине или основанию. чешуйчатой чешуи, леммы или палеи, а точка прикрепления является диагностическим признаком.Некоторые ости закручены по спирали, многие закручиваются и раскручиваются в ответ на изменения влажности и вызывающие основу леммы (часто с острой мозолью) проникать в почву или шкуры. Такие ости делятся на две части, относительно толстая колонка и концевой, тонкая щетина . Очень тонкая ость часто называют щетиной и очень короткой концевой остью mucro .
УменьшениеМожно уменьшить как количество, так и размер частей колосков.Количество цветков в колоске от одного до 20, а у некоторых трав — колоски. редуцируются только до чешуек.
Если отсутствует только яичник, то цветочек обозначается как тычинок или . мужской , хотя его также часто называют бесплодным или бесплодным. Цветочек без тычинки или завязь, с леммой и палеей или только с леммой, называются стерильными или средний , хотя также используются термины бесплодный или пустой. Цветочек с яичником описывается как фертильная .Термины бесплодный, плодородный и т. Д. могут применяться как квалификаторы к леммам соответствующих цветочков.
Хотя соцветия травы обычно обоеполые, однополые цветки (оба — мужские и самка) не редкость. Они могут встречаться в колосках с обоеполыми цветочками, в однополых колосках или в соцветиях с колосками только одного пола.
Размер чешуек, лемм и палеасов также может быть уменьшен. В Microlaena stipoides (Плакучая трава) чешуйки уменьшены до крошечных чешуек, а в Лолиум sp.(Ryegrass) одна чешуйка отсутствует на всех, кроме концевого колоска. Стерильные леммы некоторых Phalaris spp. может иметь длину 1 мм.
Другие вариантыТо, как колоски распадаются при созревании, может быть полезным диагностическим средством. характерная черта. На рахилле над чешуйками может присутствовать сустав, позволяющий соцветия опадают в зрелом возрасте, оставляя чешуйки на цветоножке. Альтернативно чешуйки могут опускаться вместе с колоском, и в этом случае говорят, что колоск расчленено ниже чешуек.Дезартикуляция также может происходить в суставы рахиллы и рахиса. Парные колоски трав, принадлежащих к трибе Andropogoneae обычно попадают как единое целое вместе с частью рахис кисти, к которой они прикреплены.
Рахилла продолжается за верхний цветочек в виде небольшой щетинки в какие-то тучные колоски. Это «удлинение» рахиллы, часто трудное видеть, полезно при идентификации.
Также важно положение редуцированных цветков в колоске.Уменьшенные соцветия может встречаться на рахилле выше или ниже плодородных цветков.
Некоторые из упомянутых вариаций, как правило, встречаются вместе и характеризуют две основные группы трав. У тухлой травы от одного до нескольких цветков на колоске. с редуцированными цветками над плодовитыми цветками и расчленением над чешуйками. Паникоидные травы демонстрируют то, что часто называют «редуцированным» рисунком колосков. У этих трав количество цветков сокращено до двух, при стерильных (уменьшенных) цветочек ниже плодовитого цветочка на очень короткой рахилле, и расчленение находится ниже чешуек.Конечно, есть много исключений из этих обобщений.
Верх
MSD1 регулирует фертильность колосков на ножке сорго посредством пути жасмоновой кислоты
Sreenivasulu, N. & Schnurbusch, T. Генетическая площадка для увеличения количества зерен в злаках. Trends Plant Sci. 17 , 91–101 (2012).
CAS Статья PubMed Google ученый
Ричардс Р.A. Выбираемые признаки для увеличения фотосинтеза сельскохозяйственных культур и урожайности зерновых культур. J. Exp. Бот. 51 (спец. №), 447–458 (2000).
CAS Статья PubMed Google ученый
Swigonova, Z. et al. Тесное разделение предков генома сорго и кукурузы. Genome Res. 14 , 1916–1923 (2004).
CAS Статья PubMed PubMed Central Google ученый
Хьюз, Т. Е., Лэнгдейл, Дж. А. и Келли, С. Влияние широко распространенной регуляторной неофункционализации на эволюцию генов гомеологов после дупликации всего генома у кукурузы. Genome Res. 24 , 1348–1355 (2014).
CAS Статья PubMed PubMed Central Google ученый
Патерсон, А. Х. и др. Геном Sorghum bicolor и разнообразие трав. Nature 457 , 551–556 (2009).
ADS CAS Статья PubMed Google ученый
Jiao, Y. et al. Улучшенный эталонный геном кукурузы с использованием одномолекулярных технологий. Природа 546 , 524–527 (2017).
ADS CAS PubMed Google ученый
Boyles, R.E. et al. Полногеномные ассоциативные исследования компонентов урожайности зерна в разнообразной зародышевой плазме сорго. Геном растений 9 (2016).
Боммерт П., Сато-Нагасава Н., Джексон Д. и Хирано Х. Ю. Генетика и эволюция соцветий и развитие цветков у трав. Physiol растительных клеток. 46 , 69–78 (2005).
CAS Статья PubMed Google ученый
Brown, P. J. et al. Наследование архитектуры соцветий у сорго. Теор.Прил. Genet. 113 , 931–942 (2006).
CAS Статья PubMed Google ученый
Burow, G., Xin, Z., Hayes, C. & Burke, J. Характеристика мутанта сорго с многосеями ( msd ) для увеличения урожайности зерна. Crop Sci. 54 , 2030–2037 (2014).
Артикул Google ученый
Дальберг, Дж. А. в Сорго: происхождение, история, технология и производство (ред. Смит, К. А. и Фредериксен, Р. А.) 99–130 (John Wiley & Sons, New York, 2000).
Wasternack, C. & Hause, B. Jasmonates: биосинтез, восприятие, передача сигнала и действие в ответ на стресс, рост и развитие растений. Обновление обзора 2007 года в Annals of Botany. Ann. Бот. 111 , 1021–1058 (2013).
CAS Статья PubMed PubMed Central Google ученый
Юань, З. и Чжан, Д. Роль передачи сигналов жасмоната в соцветии растений и развитии цветов. Curr. Opin. Plant Biol. 27 , 44–51 (2015).
CAS Статья PubMed Google ученый
Li, C. et al. Ген супрессора просистемин-опосредованных ответов2 в томате кодирует десатуразу жирных кислот, необходимую для биосинтеза жасмоновой кислоты и выработки системного раневого сигнала для экспрессии защитного гена. Plant Cell Online 15 , 1646–1661 (2003).
CAS Статья Google ученый
Лайонс, Р., Маннерс, Дж. М. и Казан, К. Биосинтез и передача сигналов жасмоната у однодольных: сравнительный обзор. Plant Cell Rep. 32 , 815–827 (2013).
CAS Статья PubMed Google ученый
Browse, J.Возможности мутантов для исследования биосинтеза и передачи сигналов жасмоната. Фитохимия 70 , 1539–1546 (2009).
CAS Статья PubMed Google ученый
Goetz, S. et al. Роль цис-12-оксофитодиеновой кислоты в развитии зародышей томатов. Plant Physiol. 158 , 1715–1727 (2012).
CAS Статья PubMed PubMed Central Google ученый
Li, L. et al. Гомолог CORONATINE-INSENSITIVE1 томатов необходим для материнского контроля созревания семян, защитных реакций, сигнализируемых жасмонатом, и развития железистых трихом. Растительная клетка 16 , 126–143 (2004).
CAS Статья PubMed PubMed Central Google ученый
Cai, Q. et al. Жасмоновая кислота регулирует развитие колосков в рисе. Nat. Commun. 5 , 3476 (2014).
PubMed Google ученый
Acosta, I. F. et al. tasselseed1 — это липоксигеназа, влияющая на передачу сигналов жасмоновой кислоты при определении пола кукурузы. Наука 323 , 262–265 (2009).
CAS Статья PubMed Google ученый
ДеЛонг, А., Кальдерон-Урреа, А. и Деллапорта, С. Л. Ген детерминации пола TASSELSEED2 кукурузы кодирует короткоцепочечную алкогольдегидрогеназу, необходимую для аборта органов цветков в зависимости от стадии. Cell 74 , 757–768 (1993).
CAS Статья PubMed Google ученый
Yan, Y. et al. Нарушение OPR7 и OPR8 раскрывает разнообразные функции жасмоновой кислоты в развитии и защите кукурузы. Растительная клетка 24 , 1420–1436 (2012).
CAS Статья PubMed PubMed Central Google ученый
Doebley, J., Stec, A. & Hubbard, L. Эволюция апикального доминирования у кукурузы. Nature 386 , 485–488 (1997).
ADS CAS Статья PubMed Google ученый
Kosugi, S. & Ohashi, Y. PCF1 и PCF2 специфически связываются с цис-элементами в гене ядерного антигена пролиферирующих клеток риса. Растительная клетка 9 , 1607–1619 (1997).
CAS Статья PubMed PubMed Central Google ученый
Луо Д., Карпентер Р., Винсент К., Копси Л. и Коэн Е. Происхождение цветочной асимметрии у Antirrhinum. Nature 383 , 794–799 (1996).
ADS CAS Статья PubMed Google ученый
Манассеро, Н. Г. У., Виола, И. Л., Велчен, Э. и Гонсалес, Д. Х. Факторы транскрипции TCP: архитектура формы растений. Biomol. Концепции 4 , 111–127 (2013).
CAS Статья PubMed Google ученый
Мартин-Трилло, М. и Кубас, П. Гены TCP: семейный снимок десять лет спустя. Trends Plant Sci. 15 , 31–39 (2010).
CAS Статья PubMed Google ученый
Francis, A. et al. Сравнительный филогеномный анализ дает представление о функциях генов TCP у сорго. Sci. Отчет 6 , 38488 (2016).
ADS CAS Статья PubMed PubMed Central Google ученый
Jiao, Y. et al. Ресурс мутанта сорго как эффективная платформа для открытия генов в травах. Растительная клетка 28 , 1551–1562 (2016).
CAS PubMed PubMed Central Google ученый
Morris, G.P. et al. Популяционно-геномные и полногеномные ассоциации агроклиматических признаков сорго. Proc. Natl. Акад. Sci. США 110 , 453–458 (2013).
ADS CAS Статья PubMed Google ученый
Mace, E. S. et al. Полногеномное секвенирование показывает неиспользованный генетический потенциал сорго, выращиваемого в Африке. Nat. Commun. 4 , 2320 (2013).
PubMed PubMed Central Google ученый
Ядзава Т., Кавахигаши Х., Мацумото Т. и Мизуно Х. Одновременный анализ транскриптома сорго и Bipolaris sorghicola с использованием RNA-seq в сочетании с сборкой транскриптома de novo. PLoS ONE 8 , e62460 (2013).
ADS CAS Статья PubMed PubMed Central Google ученый
Koppolu, R. et al. Шестирядный колос 4 (Vrs4) контролирует детерминированность колосков и тип рядков ячменя. Proc. Natl. Акад. Sci. США 110 , 13198–13203 (2013).
ADS CAS Статья PubMed PubMed Central Google ученый
Komatsuda, T. et al. Шестирядный ячмень возник в результате мутации в гене гомеобокса I-класса гомеодомен-лейциновая застежка-молния. Proc. Natl. Акад. Sci. США 104 , 1424–1429 (2007).
ADS CAS Статья PubMed PubMed Central Google ученый
Келлог, Э. А. Цветочные проявления: генетический контроль соцветий травы. Curr. Opin. Plant Biol. 10 , 26–31 (2007).
CAS Статья PubMed Google ученый
Takeda, T. et al. Ген OsTB1 отрицательно регулирует латеральное ветвление у риса. Plant J. 33 , 513–520 (2003).
CAS Статья PubMed Google ученый
Кебром, Т. Х., Берсон, Б. Л. и Финлейсон, С. А. Фитохром B подавляет экспрессию Teosinte Branched1 и индуцирует разрастание подмышечных зачатков сорго в ответ на световые сигналы. Plant Physiol. 140 , 1109–1117 (2006).
CAS Статья PubMed PubMed Central Google ученый
Youssef, H. M. et al. VRS2 регулирует опосредованное гормонами формирование рисунка соцветий ячменя. Nat. Genet. 49 , 157–161 (2017).
CAS Статья PubMed Google ученый
Рамзи, Л.и другие. INTERMEDIUM-C, модификатор фертильности боковых колосков ячменя, является ортологом гена одомашнивания кукурузы TEOSINTE BRANCHED 1. Nat. Genet. 43 , 169–172 (2011).
CAS Статья PubMed Google ученый
Bull, H. et al. Ячмень SIX-ROWED SPIKE3 кодирует предполагаемую деметилазу h4K9me2 / me3 Jumonji C-типа, которая подавляет фертильность латеральных колосков. Nat. Commun. 8 , 936 (2017).
ADS Статья PubMed PubMed Central Google ученый
Schommer, C. et al. Контроль биосинтеза и старения жасмоната мишенями miR319. PLoS Biol. 6 , e230 (2008).
Артикул PubMed PubMed Central Google ученый
Koch, J. R. et al. Чувствительность к озону гибридного тополя коррелирует с нечувствительностью как к салициловой, так и к жасмоновой кислотам.Роль запрограммированной гибели клеток в образовании повреждений. Plant Physiol. 123 , 487–496 (2000).
CAS Статья PubMed PubMed Central Google ученый
Reinbothe, C., Springer, A., Samol, I. & Reinbothe, S. Оксилипины растений: роль жасмоновой кислоты во время запрограммированной гибели клеток, защиты и старения листьев. FEBS J. 276 , 4666–4681 (2009).
CAS Статья PubMed Google ученый
Хундертмарк, М. и Хинча, Д. К. Белки LEA (поздний эмбриогенез в изобилии) и их кодирующие гены в Arabidopsis thaliana . BMC Genomics 9 , 118 (2008).
Артикул PubMed PubMed Central Google ученый
Манфре, А.Дж., Ланни, Л.М. и Маркотт, В.Р. Младший. Группа 1 Arabidopsis ПОЗДНИЙ ЭМБРИОГЕНЕЗ Изобилие белка ATEM6 требуется для нормального развития семян. Plant Physiol. 140 , 140–149 (2006).
CAS Статья PubMed PubMed Central Google ученый
Gábrišová, D. et al. Есть ли у купинов не только запасные белки для семян? Фронт. Plant Sci. 6 , 1215 (2015).
PubMed Google ученый
Duchrow, M., Hasemeyer, S., Broll, R., Брух, Х. П. и Виндховел, У. Оценка пролиферативной активности колоректальных карцином с помощью количественной полимеразной цепной реакции с обратной транскриптазой (ОТ-ПЦР). Cancer Invest. 19 , 588–596 (2001).
CAS Статья PubMed Google ученый
Xin, Z. & Chen, J. Метод высокопроизводительной экстракции ДНК с высоким выходом и качеством. Растительные методы 8 , 26 (2012).
CAS Статья PubMed PubMed Central Google ученый
Langmead, B. & Salzberg, S. L. Быстрое выравнивание по пробелам и чтению с Bowtie 2. Nat. Методы 9 , 357–359 (2012).
CAS Статья PubMed PubMed Central Google ученый
Li, H. et al. Формат Sequence Alignment / Map и SAMtools. Биоинформатика 25 , 2078–2079 (2009).
Артикул PubMed PubMed Central Google ученый
McLaren, W. et al. Получение последствий геномных вариантов с помощью Ensembl API и предиктора эффекта SNP. Биоинформатика 26 , 2069–2070 (2010).
CAS Статья PubMed PubMed Central Google ученый
Greene, E.A. et al. Спектр химически индуцированных мутаций в результате крупномасштабного обратного генетического скрининга Arabidopsis. Генетика 164 , 731–740 (2003).
CAS PubMed PubMed Central Google ученый
Coen, E. S. et al. Floricaula: гомеотический ген, необходимый для развития цветков Antirrhinum majus. Cell 63 , 1311–1322 (1990).
CAS Статья PubMed Google ученый
Jackson, D., Veit, B. & Hake, S. Экспрессия генов гомеобокса, связанных с KNOTTED1 кукурузы, в апикальной меристеме побега позволяет прогнозировать паттерны морфогенеза вегетативного побега. Разработка 120 , 405–413 (1994).
CAS Google ученый
Szklarczyk, D. et al. STRINGv10: сети белок-белкового взаимодействия, интегрированные в древо жизни. Nucleic Acids Res. 43 , D447 – D452 (2015).
CAS Статья PubMed Google ученый
Dobin, A. et al. STAR: сверхбыстрый универсальный выравниватель RNA-seq. Биоинформатика 29 , 15–21 (2013).
CAS Статья PubMed Google ученый
Trapnell, C. et al. Анализ дифференциальной экспрессии генов и транскриптов в экспериментах с последовательностью РНК с TopHat и Cufflinks. Nat. Protoc. 7 , 562–578 (2012).
CAS Статья PubMed PubMed Central Google ученый
Du, Z., Zhou, X., Ling, Y., Zhang, Z. & Su, Z. agriGO: набор аналитических инструментов GO для сельскохозяйственного сообщества. Nucleic Acids Res. 38 , W64 – W70 (2010).
CAS Статья PubMed PubMed Central Google ученый
Шмельц, Э. А., Энгельберт, Дж., Тумлинсон, Дж. Х., Блок, А. и Алборн, Х. Т. Использование парофазной экстракции в метаболическом профилировании фитогормонов и других метаболитов. Plant J. 39 , 790–808 (2004).
CAS Статья PubMed Google ученый
Границы | OsVIL2 регулирует развитие колосков путем контроля регулирующих генов у Oryza sativa
Введение
Травы имеют уникальную единицу соцветий, колоск, который содержит разное количество цветков и чешуек в зависимости от вида (Bommert et al., 2005). Колосок риса ( Oryza sativa ) состоит из двух рудиментарных чешуек, двух пустых чешуек и одного цветочка (Bommert et al., 2005). Каждый цветочек состоит из леммы, палеа, двух лодикул со стороны палеи, шести тычинок и плодолистика (Yoshida, Nagato, 2011).
Гены семействаAPETALA2 / этилен-чувствительный фактор (AP2 / ERF), SUPERNUMERARY BRACT ( SNB ) и INDETERMINATE SPIKELET1 ( IDS1 ), играют решающую роль в переходе от меристемы (SM) цветков. (FM) (Ли и др., 2007; Ли и Ан, 2012). Их мутантные линии производят повторяющиеся чешуйки и демонстрируют аномальный рисунок цветочных органов из-за расширенной активности SM. Другой ген AP2 / ERF, MULTI-FLORET SPIKELET1 ( MFS1 ), также играет роль в регуляции судьбы SM. У мутантов mfs1 появляются дополнительные леммоподобные органы и удлиненные рахиллы, а пустые чешуйки и палеа дегенерируют (Ren et al., 2013). Эти результаты предполагают, что правильный переход от SM к FM необходим для нормального развития колосков.
Анализ различных мутантов риса выявил несколько генов, участвующих в развитии чешуек. Например, мутации в EXTRA GLUME1 ( EG1 ) и EG2 / OsJAZ1 , которые функционируют в передаче сигналов жасмоновой кислоты, вызывают аномальные фенотипы колосков. Пустые чешуйки трансформируются в органы, похожие на чешуйки, и образуются лишние чешуйки (Li et al., 2009; Cai et al., 2014). Кроме того, это влияет на идентичность и количество цветочных органов. Эти изменения, вероятно, связаны с измененной экспрессией OsMADS1 у мутантов (Jeon et al., 2000а; Прасада и др., 2005). Сходные фенотипы наблюдаются для растений риса с РНКи INDETERMINATE GAMETOPHYTE1 ( OsIG1 ) (Zhang et al., 2015). Мутации в длинной стерильной лемме ( G1 ) связаны с гомеотическим преобразованием стерильной леммы в лемму, предполагая, что ген репрессирует идентичность леммы, чтобы специфицировать стерильную лемму (Yoshida et al., 2009). Мутации OsMADS34 вызывают плейотропные эффекты, включая изменение пустых чешуек в леммоподобные органы (Gao et al., 2010).
Развитие палеа задерживается у мутантов, дефектных по RETARDED PALEA1 ( REP1 ) (Yuan et al., 2009). DEPRESSED PALEA 1 ( DP1 ), кодирующий ДНК-связывающий белок AT-hook, также играет критическую роль в развитии палеа (Jin et al., 2011). Мутации этого гена вызывают дефект палеа, а также увеличение количества цветочных органов. Анализ экспрессии показал, что DP1 функционирует выше REP1 .Мутации OsMADS15 также приводят к дефектному палеа (Wang et al., 2010), тогда как мутации OsMADS6 также нарушают палею и изменяют развитие плодолистика (Ohmori et al., 2009; Li et al., 2010). Мутации OsMADS32 связаны с дефектными краевыми областями палеа и эктопических органов цветка (Sang et al., 2012).
Белки группы Polycomb (PcG) являются эпигенетическими репрессорами, которые контролируют экспрессию генов (Mozgova and Hennig, 2015). Они участвуют в различных процессах развития, образуя репрессивный комплекс Polycomb 2 (PRC2), который ингибирует целевые хроматины посредством триметилирования гистона 3, лизина 27 (h4K27me3) (Cao et al., 2002; Чермин и др., 2002; Мюллер и др., 2002; Некрасов и др., 2005). PRC2 контролирует инициирование FM, спецификацию идентичности органа и завершение меристемы (Gan et al., 2013).
PRC2 состоит из нескольких компонентов. В Arabidopsis мутанты основных компонентов PRC2 — CLF / SWN, FIE, EMF2 и MSI — представляют эктопическую экспрессию AGAMOUS ( AG ), вызывая патологические фенотипы цветков, аналогичные фенотипам AG с повышенной экспрессией. растения (Goodrich et al., 1997; Йошида и др., 2001; Хенниг и др., 2003; Moon et al., 2003; Кац и др., 2004). Этот комплекс также влияет на терминацию FM, регулируя временную экспрессию WUSCHEL ( WUS ) и KNUCKLES ( KNU ) (Mozgova et al., 2015). После того, как все органы цветков инициированы, FM завершается посредством репрессии WUS , гена поддержания меристемы (Mayer et al., 1998). PRC2 подавляет экспрессию KNU , что ингибирует транскрипцию WUS (Lenhard et al., 2001; Sun et al., 2009). Для терминации цветков активированный AG вытесняет PRC2 из KNU , что приводит к активации KNU и репрессии WUS (Sun et al., 2014).
INSENSITIVE 3 (VIN3), другой компонент PRC2, усиливает h4K27me3 в FLOWERING LOCUS C ( FLC ), репрессоре цветения (Sung and Amasino, 2004). У риса VIN3-подобный белок OsVIL2 усиливает цветение, опосредуя h4K27me3 на хроматине LFL1 , который является негативным регулятором цветения (Yang et al., 2013). OsVIL2 связывается с OsEMF2b, ортологом Arabidopsis EMF2 (Yang et al., 2013). Мутанты в OsEMF2b демонстрируют фенотипы тяжелых дефектов органов цветка и неопределенности меристемы, аналогичные мутантам, дефектным по генам E-функции OsMADS1, OsMADS6 и OsMADS34 (Luo et al., 2009; Yang et al., 2013 ; Conrad et al., 2014; Xie et al., 2015). OsEMF2b подавляет экспрессию этих генов E-функции, изменяя h4K27me3 на их хроматинах (Conrad et al., 2014). Кроме того, OsEMF2b ингибирует OsLFL1 и OsMADS4 , опосредуя h4K27me3 на их хроматине, что приводит к стимулированию цветения и регулирует спецификацию идентичности органов цветка (Xie et al., 2015).
Мутанты osvil2 также обнаруживают аномальное развитие колосков. В этом исследовании мы изучили мутантные фенотипы, которые появляются на ранних стадиях этого процесса, и провели анализ транскриптома, чтобы идентифицировать нижестоящие гены, контролируемые OsVIL2 .
Материалы и методы
Растительные материалы и условия роста
Две мутантные линии OsVIL2 , osvil2-1 и osvil2-2 , были выделены из пула линий мечения Т-ДНК риса (Jeon et al., 2000b; Jeong et al., 2002; Yang et al. ., 2013). Семена мутантов и семян дикого типа (WT) проращивали на среде MS, и было проведено генотипирование для идентификации гомозиготных растений, как объяснялось ранее (Yi and An, 2013). Растения выращивали либо на рисовом поле в естественных условиях, либо в теплице при дополнительном искусственном освещении.
Построение векторов и преобразование риса
Для конструкции слияния промотор OsVIL2 — GUS мы использовали фрагмент промотора длиной 2348 п.н. между -2340 и +8 от сайта инициации трансляции OsVIL2 , который использовали для комплементации мутанта (Zhao et al., 2010). Фрагмент амплифицировали с помощью ПЦР и помещали выше гена GUS без промотора с использованием бинарного вектора pGA3519, который содержит селектируемый маркер гигромицина.Последовательности праймеров для амплификации промоторной области перечислены в дополнительной таблице S1. Штамм LBA4404 Agrobacterium tumefaciens трансформировали этим вектором методом замораживания-оттаивания (An et al., 1989). Трансгенные растения, экспрессирующие репортерный ген GUS , были получены путем совместного культивирования клеток Agrobacterium с каллусами щитка, полученными из зрелых семян риса (сорт Longjin). Совместно культивированные каллусы были отобраны и регенерированы, как сообщалось ранее (Jeon et al., 1999).
Гистохимическое окрашивание GUS
Растительные ткани погружали в окрашивающий GUS раствор, содержащий 100 мМ фосфата натрия (pH 7,0), 0,1 мМ феррицианида калия, 0,1 мМ ферроцианида калия, 0,1% Triton X-100, 10 мМ EDTA (pH 8,0), 1% DMSO, 0,1% X-gluc (5-бром-4-хлор-3-индолил-β- D -глюкуроновая кислота / соль циклогексиламмония) и 5% метанола (Yoon et al., 2014). Образцы инкубировали в течение ночи при 37 ° C, затем переносили в 70% этанол при 65 ° C на несколько часов для удаления хлорофиллов перед хранением в 95% этаноле.
Сканирующая электронная микроскопия
Образцы были подготовлены, как описано ранее (Lee et al., 2007). Образцы фиксировали в растворе FAA, дегидратировали в этаноле и сушили до критической точки в Leica EM CPD300 (Leica Microsystems, Вецлар, Германия). Их устанавливали на штыри, покрывали платиной напылением и наблюдали под растровым электронным микроскопом (SIGMA FE-SEM; Carl Zeiss, Германия).
Выделение РНК и анализ qRT-PCR
тотальных РНК выделяли с помощью RNAiso Plus (TaKaRa, Shiga, Japan).КДНК были синтезированы с использованием 2 мкг РНК, обратной транскриптазы вируса мышиного лейкоза Молони (Promega, Мэдисон, Висконсин, США), ингибитора рибонуклеазы RNasin (Promega), 10 нг праймера олиго (dT) 18 и 2,5 мМ дезоксирибонуклеотидтрифосфаты. Количественная ОТ-ПЦР была проведена с использованием SYBR Green I Prime Q-Mastermix (GENETBIO, Тэджон, Южная Корея) и Rotor-Gene 6000 (Corbett Research, Сидней, Новый Южный Уэльс, Австралия) в соответствии с протоколами, описанными ранее (Yang et al., 2013 ; Wei et al., 2016). Рис Убиквитин 1 использовали в качестве внутреннего контроля для количественной ПЦР в реальном времени (qRT-PCR).
Для транскриптомных анализов суммарные РНК получали из 2- и 4-мм метелок растений WT и osvil2-1 в трех биологических повторностях. Двухцепочечные кДНК синтезировали со случайными гексамерами и лигировали с адаптерами. Эта библиотека была секвенирована попарно с использованием стратегии PE90 на Illumina HiSeq TM 2000 в Пекинском институте геномики (Ухань, Китай).Алгоритм DEseq применялся для фильтрации дифференциально экспрессируемых генов. Все аннотации Gene Ontology (GO) были загружены с NCBI, UniProt и веб-сайта Gene Ontology. Данные о секвенировании РНК были депонированы в базу данных GEO (номер доступа: GSE108538).
Результаты
Мутации в
OsVIL2 , вызвавшие аномальное образование колосковРанее мы показали, что мутантные линии с инсерцией Т-ДНК osvil2-1 и osvil2-2 демонстрируют плейотропные фенотипы, включая позднее цветение, меньшее количество побегов и аномальное развитие колосков (Yang et al., 2013). В этом исследовании мы подробно охарактеризовали дефекты колосков (рисунок 1 и дополнительный рисунок S1). Колоски WT имели пару рудиментарных чешуек плюс пустые чешуйки (рис. 1А). Напротив, количество рудиментарных чешуек было увеличено в 14% колосков osvil2 (рис. 2А). Кроме того, пустые чешуи были удлинены у 21% мутантных колосков (Рисунок 1B и дополнительный рисунок S1B), в то время как количество пустых чешуек было уменьшено на 21%, в результате чего колоски были с одной пустой чешуей или без нее (Рисунки 1C, 2B) .Иногда на месте пустой чешуйки развивался леммоподобный орган. Эти наблюдения показали, что мутации OsVIL2 влияют как на количество, так и на морфологию пустых чешуек.
РИСУНОК 1. Фенотип Spikelet WT и мутант osvil 2. (A – E) Фенотипы WT и osvil2-1 колосков. (A) Колосок WT с парой рудиментарных чешуек и пустой чешуей, лемма, палеа. (Б) osvil2-1 колоск с удлиненными пустыми чешуйками. (C) osvil2-1 колоск с дегенерированной палеей и без пустой чешуи на стороне леммы (стрелка). (D) osvil2-1 колоск с дополнительным леммоподобным органом. (E) osvil2-1 колоск с 2 леммами и 2 вырожденными палеа. (F – K) Фенотипы внутренних органов цветков WT и osvil2-1 . Палеа и лемма были удалены. (F) Колоск WT состоит из двух лодикул на стороне леммы, шести тычинок (звездочек) и одного пестика с двумя рыльцами (стрелки). (G) osvil2-1 цветочек с лишними лодикулами и незрелыми тычинками. (H) osvil2-1 цветочек с мозаичными органами lodicule – тычинки. (I) osvil2-1 цветочек с лишними лодикулами, семью тычинками (звездочками) и двумя плодолистиками. (J) osvil2-1 плодолистик, в котором слито несколько плодолистиков. Количество клейм также увеличено (стрелки). (K) Мозаичный орган Lodicule – тычинки в osvil2-1 . (L) Поперечное сечение колоска WT. (M, N) Поперечное сечение колоса osvil2 . (М) Образование дополнительного леммоподобного органа в колоске osvil2 . (N) osvil2 с двумя леммами, двумя дегенерированными палеа, тремя аномальными лодикулами (наконечниками стрелок) и одним сросшимся плодолистиком с двумя семязачатками. боп, тело палеа; Ca, плодолистник; DP — депрессивная палеа; EG, пустая чешуйка; ЭЭГ — удлиненная пустая чешуйка; L, лемма; Lo, lodicule; L / S — мозаичный орган lodicule – тычинка; Мне нравится; леммоподобный орган; mrp — краевая область палеи; Ov, яйцеклетка; P, palea; RG, рудиментарная чешуйка.Звездочками обозначены тычинки (F, I) или сосудистые пучки (L – N) . Масштабная линейка = 1 мм (A – K) или 500 мкм (L – N) .
РИСУНОК 2. Количество цветочных органов в WT, osvil2-1 и osvil2-2 колосков. (A) Рудиментарные чешуйки; (B) пустых чешуек; (C) лемма и палеа; (D) lodicules; (E) тычинок; (F) пестиков.
Кроме того, у osvil2 соцветий было аномально развитие всех органов цветков.Дополнительные леммоподобные структуры наблюдались в 45% колосков osvil2 , создавая три или более таких структур (рисунки 1D, 2C и дополнительный рисунок S1C). Дополнительные леммоподобные органы часто образовывались на эктопическом обороте. Некоторые из этих эктопических леммоподобных органов напоминали лемму (рис. 1D). Количество лемм увеличилось в 14% колосков из osvil2 , и они часто сопровождали дегенерированную палею (Рисунки 1E, 2C). Развитие палеа было дефектным у 33% этих мутантов (рисунок 1C и дополнительный рисунок S1D), а поперечные сечения колосков показали дополнительные леммоподобные органы (рисунок 1M) и дегенерированный палеа (рисунок 1N).
Число лодикул увеличилось до трех или более у 54% колосков osvil2 (Рисунки 1G – I, 2D и дополнительные рисунки S1F, G), и их морфология иногда была аномальной. Лодикулы были удлиненными (20%) или похожим на пыльник органом, образованным в верхней части 26% этих лодикул (Рисунки 1 H, I, K). Количество тычинок уменьшилось у 8% колосков (Рисунки 1H, 2E), но увеличилось в 22% (Рисунки 1I, 2E). Наконец, количество плодолистиков увеличилось до двух у 32% мутантных колосков (Рисунки 1I, 2F), и они часто сливались (Рисунки 1 H, J).Эти наблюдения показали, что OsVIL2 необходим для правильного развития всех органов в колоске.
Раннее развитие колосков было затронуто в
osvil2Мы использовали растровый электронный микроскоп для изучения дефектов колосков на ранних стадиях развития, которые были классифицированы на основании предыдущих исследований (Ikeda et al., 2004). У WT FM продуцировал пустую чешуйку и лемму на противоположной стороне от пустой чешуи во время стадии развития колосков Sp3 в WT (Figure 3A).Впоследствии Palea развивалась на стадиях 4–5 (рис. 3B). В то время как лемма и палеа удлинялись, тычинки развились на стадии 6 (рис. 3C). Наконец, плодолистик вырос в центре колоска на стадии 8 (рис. 3D).
РИСУНОК 3. Сканирующие электронные микрографические изображения колосков WT и osvil2 на ранних стадиях развития. (A – D) колосков WT. (A) Этап Sp3 во время формирования зачатков леммы. (B) Стадия Sp4-5 формирования зачатков палеа; возможно, внутри леммы зарождаются два зачатка лодикулы. (C) Стадия Sp6 с образованием шести зачатков тычинок (звездочка). (D) Стадия Sp8, с зарождением зачатков плодолистиков во время роста леммы и палеи. (E – H) Замедленный рост палеа у osvil2 колосков. (E) Стадия Sp4, демонстрирующая замедление роста зачатков палеа (стрелка) по сравнению с ростом леммы. (F) Стадия Sp6 с дегенеративным формированием палеа и замедленным формированием зачатков тычинок на палеа. (G) Стадия Sp8, с замедленным ростом палеи по сравнению с леммой. (H) Стадия Sp6, демонстрирующая неправильное формирование зачатков тычинок (звездочка). Дополнительный зачаток органа (наконечник стрелки) между палеей и тычинкой, увеличивающий размер цветочного мутовки. (I – L) osvil2 колоск с фенотипом двойных цветков. (I) Стадия Sp3, с более широкой цветочной меристемой и двумя зачатками лемм с обеих сторон. (J) Стадия Sp4, с двумя вырожденными зачатками палеи, образующимися в середине двух лемм. (K) Стадия Sp6, представляющая зачатки эктопических органов (наконечники стрелок), вероятно, образующие дополнительные палеа и лодикулы. (L) Stage Sp6-7, с образованием восьми зачатков тычинок. (M – P) osvil2 колоск с дополнительной чешуей или леммоподобными органами. (M) Стадия Sp7, на которой количество зачатков тычинок снижено до 5. Цветочные мутовки увеличиваются, и после дополнительной леммы формируется дегенерированная палеа. (N) Стадия Sp4 с образованием дополнительных зачатков чешуек (стрелки). (O) Стадия Sp6, на которой производятся дополнительные леммы, в то время как формирование внутреннего органа задерживается. (P) Стадия Sp8, с дополнительной леммой, полученной после образования нормальных пустых чешуек и рудиментарных чешуек. Ca, плодолистник; DP — дегенерированная палеа; EG, пустая чешуйка; FM — цветочная меристема; L, лемма; P, palea; RG, рудиментарная чешуйка. Масштабные линейки = 20 мкм.
У osvil2 колосков образование палеа зачатков часто задерживалось на Sp4 (рис. 3E). На Sp6 мутантные колоски обнаруживают дегенерированные палеа-зачатки, часто вместе с задержкой развития внутренних органов цветков на палеа-стороне (Рисунки 3F, G).Это замедление, по-видимому, привело к снижению количества производимых тычинок, а также к аномальному развитию внутренних органов цветка (рис. 3H). FM были больше у некоторых колосков osvil2 , которые обычно сопровождали два зачатка леммы и два дегенерированных зачатка палеа (рис. 3I, J). На Sp6 между леммой и зачатком тычинок наблюдались дополнительные зачатки лодикулов (рис. 3K). Во время этой стадии количество зачатков тычинок было изменено (Рисунки 3L, M), и повторяющееся образование чешуек иногда происходило (Рисунок 3N).Дополнительные чешуйки и леммоподобные органы образовывались на дополнительных оборотах (Рисунки 3M – P). Эти находки показали, что OsVIL2 функционирует на ранних стадиях развития колосков.
Паттерн экспрессии
OsVIL2Характер экспрессии OsVIL2 анализировали с помощью ОТ-ПЦР. В вегетативных тканях ген был обнаружен в листьях проростков и листовых пластинках зрелых растений (рис. 4А). Он также конститутивно экспрессировался в метелках на разных стадиях развития (Рис. 4B).У зрелых колосков экспрессия была сильной в тычинках и плодолистиках и слабой в лодикулах и палеа (рис. 4С). Экспрессию также изучали на тканевом уровне с использованием промоторной области OsVIL2 , слитой с репортерным геном GUS (фиг. 4D). Мы получили 13 растений, независимо трансформированных этой конструкцией промотор OsVIL2 — GUS . Эти линии отображали похожий образец выражения, поэтому мы выбрали строку с самым высоким выражением GUS для дальнейшего анализа.Репортер GUS сильно экспрессировался в листьях, но не экспрессировался в корнях, как наблюдали из анализов RT-PCR (Фигуры 4E, F). Во время развития колосков репортер был обнаружен в базальных областях колосков (Рисунки 4G – J). У цветков Sp8 он сильно выражен в пыльниках и базальной области плодолистиков (Рисунки 4K – M). Этот предпочтительный для органов паттерн экспрессии соответствовал паттерну, выявленному с помощью данных qRT-PCR.
РИСУНОК 4. Характер экспрессии OsVIL2 в различных органах. (A – C) Анализ экспрессии ОТ-ПЦР. (A) Экспрессия в различных органах. (B) Экспрессия во время раннего развития колосков. (C) Экспрессия в каждом органе цветка в зрелых колосках. (D) Принципиальная схема конструкции pOsVIL2 :: GUS . (E – M) Наблюдение за линией улавливания промотор-GUS OsVIL2. Экспрессия в листовой пластинке (E) , корень (F) , колоск на Sp4 (G) , колоск на Sp5 (H) , колоск на Sp6 (I) , колоск на Sp7 ( J) , колоски в 12-миллиметровой молодой метелке (K) , колоски в 50-миллиметровой метелке (L) и зрелом колоске (M) .EG, пустая чешуйка; L, лемма; P, palea; RG, рудиментарная чешуйка.
Анализ транскриптома молодых метелок
Были проведены анализы секвенированияРНК для идентификации генов, дифференциально экспрессируемых в osvil2 во время раннего развития колосков. Были исследованы две стадии: 2-миллиметровые метелки, содержащие колоски в основном на Sp4 или ранее, и 4-миллиметровые метелки, содержащие колоски в основном на Sp7 или ранее. В 2-миллиметровых образцах 451 ген был активирован (дополнительная таблица S2), а 606 генов подавлен (дополнительная таблица S3) по крайней мере в два раза.В 4-миллиметровых образцах 548 генов были активированы (дополнительная таблица S4), а 490 генов подавлены (дополнительная таблица S5) по крайней мере в два раза. Перекрытие между 2- и 4-миллиметровыми образцами показало, что у 330 генов была повышенная регуляция (дополнительная таблица S6), в то время как у 306 генов была понижена регуляция (дополнительная таблица S7). В общей сложности 669 генов были активированы, а 790 генов подавлены по крайней мере в два раза в обоих классах размеров (рис. 5).
РИСУНОК 5. Число генов, дифференциально экспрессируемых в osvil2 молодых метелках. (A) Число увеличено; Номер (B) с пониженным регулированием.
Мы выполнили анализ GO с использованием дифференциально экспрессируемых генов (дополнительный рисунок S2). Наш анализ GO показал, что развитие цветка (GO: 0009908) было значительно обогащено генами с пониженной регуляцией в 2-мм метелках, что означает, что регуляторные гены, участвующие в этом процессе, были подавлены в osvil2 на ранних стадиях. Термины для транскрипции (GO: 0006351) и регуляции транскрипции (GO: 0006355) также были высоко обогащены для генов с пониженной регуляцией от обоих размеров метелки (дополнительные рисунки S2B, D).Среди факторов транскрипции многие гены семейств AP2 и MADS-боксов были значительно подавлены (дополнительная таблица S3). Поскольку несколько генов внутри этих семейств играют важные функции в формировании SM и развитии цветковых органов, их подавление в развивающихся метелках, вероятно, является причиной аномальных фенотипов колосков.
Анализ транскриптомов генов, участвующих в фазовом переходе меристемы или развитии органа колосков, показан в таблице 1. Гены, определяющие размер меристемы — FON1, FON4 и OsWUS (Suzaki et al., 2004; Чу и др., 2006; Moon et al., 2006; Nardmann and Werr, 2006) — почти одинаково экспрессировались у мутантов osvil2 . Среди генов, которые функционируют при переходе от меристемы соцветия (IM) к SM, частота транскриптома APO1 была значительно снижена в обоих размерах метелки. APO1 , ортолог Arabidopsis UFO , подавляет более раннее преобразование IM в SM и способствует пролиферации клеток в IM (Ikeda et al., 2005, 2007; Ikeda-Kawakatsu et al., 2009). Кроме того, APO1 регулирует идентичность органов цветка и детерминированность цветков путем усиления экспрессии гена C-функции OsMADS3 (Ikeda et al., 2005). Однако мы обнаружили, что уровень экспрессии APO2 / RFL был повышен у наших мутантов osvil2 . Ген APO2 / RFL , ортолог Arabidopsis LFY , также подавляет переход от IM к SM. Снижение его экспрессии снижает количество продуцируемых ветвей метелки и изменяет идентичность органов цветка (Rao et al., 2008; Икеда-Кавакацу и др., 2012).
ТАБЛИЦА 1. Уровни экспрессии генов, контролирующих развитие колосков.
Среди четырех генов RCN , гомологичных Arabidopsis TERMINAL FLOWER 1 (Nakagawa et al., 2002), экспрессия RCN4 была увеличена как минимум в два раза в мутантных колосках, а экспрессия RCN1, RCN2 и RCN3 существенно не пострадали. Кроме того, мы не смогли обнаружить каких-либо значительных изменений в уровнях транскриптомов для SNB и MFS , которые функционируют во время перехода от SM к FM (Таблица 1).Экспрессия была немного увеличена для FZP , гена, который ингибирует подмышечную меристему в колосках и способствует FM за счет усиления экспрессии B-функции ( OsMADS4 и OsMADS16 ), E-функции ( OsMADS1, OsMADS7 и OsMADS8 ) и AGL6-подобных ( OsMADS6 и OsMADS17 ) гены MADS-бокса (Komatsu et al., 2003; Bai et al., 2016).
Среди генов, необходимых для образования пустых чешуек (Yoshida et al., 2009; Zhang et al., 2015), G1 и OsIG1 были значительно подавлены в молодых колосках из osvil2 . Поскольку развитие пустых чешуек было ненормальным, и их количество было меньше у мутанта, снижение экспрессии генов могло быть ответственным за эти дефекты. Более того, подавление REP1 , которое необходимо для развития палеа (Yuan et al., 2009), может быть связано с дегенерацией палеа у osvil2 .
Несколько генов MADS-box, которые функционируют в идентичности органов цветка, были подавлены в колосках osvil2 .Экспрессия B-функции OsMADS4 и OsMADS16 была затронута у мутанта, особенно в 2-мм колосках. C-функция OsMADS3, OsMADS58 и DL также подавлялась в мутантных колосках. Кроме того, экспрессия была значительно снижена для D-функции OsMADS13 и E-функции OsMADS1, OsMADS6, OsMADS7 и OsMADS8 . Напротив, экспрессия генов A-функции OsMADS14, OsMADS15 и OsMADS18 существенно не пострадала, в то время как экспрессия OsMADS5 была увеличена.
Чтобы проверить наши данные о секвенировании РНК, мы провели qRT-PCR, сосредоточив внимание на 13 генах MADS-бокса, которые контролируют идентичность и развитие органов цветка (рис. 6). Среди них восемь ( OsMADS1, OsMADS3, OsMADS4, OsMADS6, OsMADS7, OsMADS13, OsMADS16 и OsMADS58 ) были подавлены, в то время как экспрессия пяти ( OsMADS2, OsMADS145, OsMADS 902, OsMADS 902, OsMADS 902, OsMADS 902, OsMADS 902 и OsMADS 902 ) не претерпел значительных изменений, согласно анализам транскриптома (рис. 6В).В мутантах osvil2 экспрессия всех восьми подавленных генов была значительно снижена на основании результатов qRT-PCR. Среди пяти, для которых анализ РНК-секвенирования не показал значительных изменений, экспрессия была сходной между WT и osvil2 для трех, в то время как два других, OsMADS5 и OsMADS34 , были слегка повышены в мутантных метелках.
РИСУНОК 6. Проверка результатов секвенирования РНК с помощью qRT-PCR.Уровни транскрипции относятся к Ubi1 . (A) Анализ qRT-PCR генов, участвующих в развитии колосков. (B) Анализ qRT-PCR генов MADS-бокса, участвующих в развитии органов цветка. (C) Анализ qRT-PCR активированных генов, ранее идентифицированных в анализе секвенирования РНК.
Мы также отобрали восемь генов, которые, как показали наши анализы транскриптомов, имеют повышенную регуляцию (рис. 6C). Эксперименты по проверке qRT-PCR показали, что их экспрессия была намного выше у мутантов osvil2 .Все эти результаты продемонстрировали надежность данных, полученных в результате анализа секвенирования РНК.
Обсуждение
В этом исследовании мы проанализировали аномальные фенотипы osvil2 и обнаружили, что они изменчивы во всех органах колосков. Более того, у мутанта была значительно изменена экспрессия основных регуляторных генов для развития колосков. Эти результаты продемонстрировали, насколько необходим OsVIL2 для формирования нормального паттерна органов во время образования колосков, поскольку он модулирует правильную экспрессию тех генов, которые контролируют развитие органов и идентичность в колосках.
Колоски мутанта osvil2 давали удлиненные пустые чешуйки, напоминающие лемму. Гомеотическая трансформация пустых чешуек в лемму также была описана для мутантов, дефектных в G1, EG1, EG2, OsMADS34 и OsIG1 (Li et al., 2009; Yoshida et al., 2009; Gao et al., 2010 ; Cai et al., 2014; Zhang et al., 2015). Поскольку пустые чешуйки не встречаются в колосках других трав, происхождение и идентичность пустых чешуек спорны (Yoshida and Nagato, 2011).Фенотип удлиненной пустой чешуи osvil2 подтверждает идею о том, что пустые чешуйки являются вырожденной леммой о стерильных соцветиях (Arber, 1934). Экспрессия генов идентичности пустой колосковой чешуи — G1 и OsIG1 — была значительно ниже в развивающихся метелках osvil2 , чем у WT, что позволяет предположить, что OsVIL2 функционирует выше G1 и OsIG1 до указать стерильное тождество леммы.
Другим фенотипом колосков osvil2 было образование дегенерированной палеи.Этот дефект имел место в основном в палеа теле, а не в краевой области. Кроме того, средняя часть зачатков палеа часто не росла, что приводило к расщеплению палеи. Эти фенотипы аналогичны фенотипам, описанным для мутантов, дефектных в REP1, OsMADS15, MFS1, DP1 или OsIG1 (Yuan et al., 2009; Wang et al., 2010; Jin et al., 2011; Ren et al. ., 2013; Zhang et al., 2015). Экспрессия REP1, DP1 и OsIG1 была снижена у мутантов osvil2 , что указывает на то, что эти гены связаны с палеа-дефектами.У Arabidopsis гены VIL функционируют во время яровизации и цветения (Sung and Amasino, 2004; Sung et al., 2006; Greb et al., 2007). Наше наблюдение, что рис VIL функционирует в развитии колосков, указывает на диверсифицированную функцию семейства генов. Поскольку колоск представляет собой уникальную структуру соцветий травы, будет интересно изучить, сохраняется ли функция OsVIL2 у других видов трав.
Наиболее значимым фенотипом osvil2 было увеличение числа всех органов цветка.Этот фенотип был подобен фенотипу мутантов, дефектных по FON4 , ортологу Arabidopsis CLV3 (Chu et al., 2006). Здесь количество цветковых органов во внутренних оборотах сильнее сказывается на колосках. Для мутантов fon4 число плодолистиков может увеличиваться до 10, тогда как у osvil2 это число возросло до двух. Число тычинок также увеличивается до 10 у fon4 по сравнению с 8 у osvil2 . Гомеотическое преобразование пустых чешуек в лемму является общим для мутантов fon4 и osvil2 .Мы отметили, что экспрессия FON4 была снижена в развивающихся метелках osvil2 , предполагая, что этот ген функционирует ниже OsVIL2 .
Идентичность цветочных органов регулируется многочисленными генами, включая некоторые гены MADS-box (Zhang and Yuan, 2014). Мы наблюдали, что несколько генов MADS-бокса по-разному экспрессировались в osvil2 . Изменение экспрессии этих цветочных гомеотических генов могло быть ответственным за аномальное развитие цветочных органов у мутанта.
Это исследование показало, что OsVIL2 влияет на различные аспекты развития колосков, контролируя различные гены, важные для развития колосков. Поскольку OsVIL2 функционирует вместе с PRC2, который подавляет хроматин-мишень, мы ожидали обнаружить, что экспрессия прямых мишеней будет выше у мутантов osvil2 . Вместо этого уровни транскрипции для большинства регуляторных генов, которые контролируют количество или идентичность органов, были снижены, тогда как экспрессия была немного увеличена для RCN4, OsMADS5 и OsMADS34 .Это подразумевает, что они могут быть прямыми мишенями OsVIL2-PRC2. Хотя функция RCN4 остается неизвестной, избыточная экспрессия RCN1 и RCN2 может приводить к сильно разветвленным метелкам, указывая тем самым, что они также играют роль в подавлении судьбы цветков (Nakagawa et al., 2002). В более раннем функциональном исследовании OsEMF2b, OsMADS34 был предсказан как прямой ген-мишень для Os EMF2b (Conrad et al., 2014). Поскольку OsEMF2b взаимодействует с OsVIL2 (Yang et al., 2013), мутации OsVIL2 также могут влиять на гены MADS-бокса.
Авторские взносы
GA организовала все это исследование. GA, HY, JY, WL и DZ разработали исследование. HY, JY, WL и DZ провели эксперименты и проанализировали данные. HY и GA написали рукопись. Все авторы прочитали и одобрили рукопись.
Финансирование
Эта работа была поддержана грантом Программы совместных исследований для развития сельскохозяйственных наук и технологий, Администрация сельского развития, Южная Корея (проект №PJ013210) в GA.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы благодарят Kyungsook An за работу с семенным фондом и Priscilla Licht за редактирование англоязычного содержания статьи.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https: // www.frontiersin.org/articles/10.3389/fpls.2018.00102/full#supplementary-material
РИСУНОК S1 | Фенотипы освил2-2 колосков. (A) Аномальное образование рудиментарной чешуи в колоске osvil2-2 . (В) osvil2-2 колоск с удлиненной пустой чешуей. (C) osvil2-2 колоск с дополнительными леммоподобными органами, обозначенными стрелками. (D) osvil2-2 колоск с дегенерированной палеей. (E) Фенотип с двумя цветками. (F) Образование дополнительных органов цветка. (G) Дополнительное образование лодикулы и удлиненное лодикуло. (H) Аномальный плодолистик с увеличенным количеством рыльц и недифференцированной клеточной массой. (I) Мозаичный орган Lodicule – тычинки в osvil2-2 . Ca, плодолистник; ЭЭГ — удлиненная пустая чешуйка; Эло, удлиненное лодикула; L, лемма; Lo, lodicule; P, palea; ucm, недифференцированная клеточная масса. Шкала шкалы = 1 мм.
РИСУНОК S2 | Генеонтологический анализ дифференциально экспрессируемых генов в osvil2 . (A) Обогащение активированных генов в 2-мм метелке. (B) Обогащение генов с пониженной регуляцией в 2-мм метелке. (C) Обогащение активированных генов в 4-мм метелке. (D) Обогащение генов с пониженной регуляцией в 4-мм метелке.
ТАБЛИЦА S1 | Последовательности праймеров, используемых в этом исследовании.
ТАБЛИЦА S2 | Гены активированы в 2-мм молодых метелках osvil2 .
ТАБЛИЦА S3 | Гены подавляются в 2-мм молодых метелках osvil2 .
ТАБЛИЦА S4 | Гены активируются в молодых 4-мм метелках osvil2 .
ТАБЛИЦА S5 | Гены подавляются в 4-мм молодых метелках osvil2 .
ТАБЛИЦА S6 | Гены активируются как в 2-, так и в 4-мм молодых метелках osvil2 .
ТАБЛИЦА S7 | Гены подавляются как в 2-, так и в 4-миллиметровых молодых метелках osvil2 .
Сноски
- http: // www.ncbi.nlm.nih.gov/
- http://www.uniprot.org/
- http://www.geneontology.org/
Список литературы
Ан, Г., Эберт, П. Р., Митра, А., и Ха, С. Б. (1989). «Бинарные векторы», в Руководство по молекулярной биологии растений , ред. С. Б. Гельвин и Р. А. Шильпероорт (Дордрехт: Kluwer Academic Publisher), 1–19.
Google Scholar
Арбер, А. (1934). Зерновые: исследование злаков, бамбука и трав .Кембридж: Издательство Кембриджского университета.
Google Scholar
Бай, X., Хуан, Ю., Мао, Д., Вэнь, М., Чжан, Л., и Син, Ю. (2016). Регуляторная роль FZP в определении ветвления метелки и образования колосков у риса. Sci. Rep. 6: 19022. DOI: 10.1038 / srep19022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Боммерт П., Сато-Нагасава Н., Джексон Д. и Хирано Х. Ю. (2005). Генетика и эволюция соцветий и развития цветков у трав. Physiol растительных клеток. 46, 69–78. DOI: 10.1093 / pcp / pci504
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цай, К., Юань, З., Чен, М., Инь, К., Ло, З., Чжао, X., и др. (2014). Жасмоновая кислота регулирует развитие колосков в рисе. Nat. Commun. 5: 3476. DOI: 10.1038 / ncomms4476
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Cao, R., Wang, L., Wang, H., Xia, L., Erdjument-Bromage, H., Tempst, P., et al.(2002). Роль метилирования гистона h4 лизина 27 в сайленсинге Polycomb-группы. Наука 298, 1039–1043. DOI: 10.1126 / science.1076997
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Chu, H., Qian, Q., Liang, W., Yin, C., Tan, H., Yao, X., et al. (2006). Ген FLORAL ORGAN NUMBER4 , кодирующий предполагаемый ортолог Arabidopsis CLAVATA3, регулирует размер апикальной меристемы у риса. Plant Physiol. 142, 1039–1052. DOI: 10.1104 / стр. 106.086736
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Конрад, Л. Дж., Хандай, И., Джонсон, К., Гвидердони, Э., Ан, Г., Виджайрагаван, У. и др. (2014). Ген EMF2B поликомбовой группы необходим для поддержания детерминированности меристемы цветков риса. Plant J. 80, 883–894. DOI: 10.1111 / tpj.12688
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чермин, Б., Мелфи, Р., МакКейб, Д., Зейтц, В., Имхоф А. и Пирротта В. (2002). Drosophila энхансер комплексов Zeste / ESC обладают активностью гистон-h4-метилтрансферазы, которая маркирует хромосомные участки Polycomb. Cell 111, 185–196. DOI: 10.1016 / S0092-8674 (02) 00975-3
CrossRef Полный текст | Google Scholar
Ган Э. С., Хуанг Дж. И Ито Т. (2013). Функциональные роли модификации гистонов, ремоделирования хроматина и микроРНК в развитии цветка Arabidopsis . Внутр. Rev. Cell Mol.Биол. 305, 115–161. DOI: 10.1016 / B978-0-12-407695-2.00003-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гао, X., Лян, W., Yin, C., Ji, S., Wang, H., Su, X., et al. (2010). Ген SEPALLATA , подобный гену OsMADS34 , необходим для развития соцветий и колосков риса. Plant Physiol. 153, 728–740. DOI: 10.1104 / стр.110.156711
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гудрич, Дж., Пуангсомли, П., Мартин, М., Лонг, Д., Мейеровиц, Э. М., и Коупленд, Г. (1997). Ген группы Polycomb регулирует экспрессию гомеотического гена в Arabidopsis . Nature 386, 44–51. DOI: 10.1038 / 386044a0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Греб Т., Майлн, Дж. С., Кревиллен, П., Джеральдо, Н., Ан, Х., Гендалл, А. Р. и др. (2007). Фингерпринтный белок VRN5 PHD действует в эпигенетическом замалчивании Arabidopsis FLC . Curr. Биол. 17, 73–78. DOI: 10.1016 / j.cub.2006.11.052
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хенниг, Л., Таранто, П., Вальзер, М., Шёнрок, Н., и Груиссем, В. (2003). Arabidopsis MSI1 необходим для эпигенетического поддержания репродуктивного развития. Разработка 130, 2555–2565. DOI: 10.1242 / dev.00470
CrossRef Полный текст | Google Scholar
Икеда, К., Ито, М., Нагасава, Н., Киодзука, Дж.и Нагато Ю. (2007). Рис ABERRANT PANICLE ORGANIZATION 1 , кодирующий белок F-бокса, регулирует судьбу меристемы. Plant J. 51, 1030–1040. DOI: 10.1111 / j.1365-313X.2007.03200.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Икеда, К., Нагасава, Н., и Нагато, Ю. (2005). ABERRANT PANICLE ORGANIZATION 1 регулирует во времени идентичность меристемы риса. Dev. Биол. 282, 349–360. DOI: 10.1016 / j.ydbio.2005.03.016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Икеда, К., Сунохара, Х., и Нагато, Ю. (2004). Ход развития соцветия и колоска у риса. Порода. Sci. 54, 147–156. DOI: 10.1270 / jsbbs.54.147
CrossRef Полный текст | Google Scholar
Икеда-Кавакацу, К., Маэкава, М., Идзава, Т., Ито, Дж. И. и Нагато, Ю. (2012). ABERRANT PANICLE ORGANIZATION 2 / RFL , ортолог риса Arabidopsis LEAFY , подавляет переход от меристемы соцветия к меристеме цветков посредством взаимодействия с APO1 . Plant J. 69, 168–180. DOI: 10.1111 / j.1365-313X.2011.04781.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Икеда-Кавакацу, К., Ясуно, Н., Оикава, Т., Иида, С., Нагато, Ю., Маекава, М., и др. (2009). Уровень экспрессии ABERRANT PANICLE ORGANIZATION1 определяет форму соцветия риса посредством контроля пролиферации клеток в меристеме. Plant Physiol. 150, 736–747. DOI: 10.1104 / стр.109.136739
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чон, Дж.С., Чунг, Ю. Ю., Ли, С., Йи, Г. Х., О, Б. Г. и Ан, Г. (1999). Выделение и характеристика гена, специфичного для пыльников, RA8, из риса ( Oryza sativa L.). Завод Мол. Биол. 39, 35–44. DOI: 10.1023 / A: 1006157603096
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Jeon, J. S., Jang, S., Lee, S., Nam, J., Kim, C., Lee, S. H., et al. (2000a). листовая шелуха стерильная1 — гомеотическая мутация в гене MADS-бокса риса, влияющая на развитие цветков риса. Растительная клетка 12, 871–884. DOI: 10.1105 / tpc.12.6.871
CrossRef Полный текст | Google Scholar
Jeon, J.S., Lee, S., Jung, K.H., Jun, S.H., Jeong, D.H., Lee, J., et al. (2000b). Инсерционный мутагенез Т-ДНК для функциональной геномики риса. Plant J. 22, 561–570. DOI: 10.1046 / j.1365-313x.2000.00767.x
CrossRef Полный текст | Google Scholar
Jeong, D.H., An, S., Kang, H.G., Moon, S., Han, J.J., Park, S., et al. (2002).Инсерционный мутагенез Т-ДНК для активации мечения в рисе. Plant Physiol. 130, 1636–1644. DOI: 10.1104 / pp.014357
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Jin, Y., Luo, Q., Tong, H., Wang, A., Cheng, Z., Tang, J., et al. (2011). Ген AT-hook необходим для формирования палеа и контроля количества цветочных органов у риса. Dev. Биол. 359, 277–288. DOI: 10.1016 / j.dbio.2011.08.023
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кац, А., Олива, М., Москна, А., Хаким, О., и Охад, Н. (2004). Поликомб белки FIE и CURLY LEAF взаимодействуют в регуляции экспрессии генов гомеобокса во время развития спорофитов. Plant J. 37, 707–719. DOI: 10.1111 / j.1365-313X.2003.01996.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Komatsu, M., Chujo, A., Nagato, Y., Shimamoto, K., and Kyozuka, J. (2003). FRIZZY PANICLE необходим для предотвращения образования подмышечных меристем и установления идентичности цветочной меристемы в рисовых колосках. Разработка 130, 3841–3850. DOI: 10.1242 / dev.00564
CrossRef Полный текст | Google Scholar
Ли, Д. Ю., и Ан, Г. (2012). Два гена семейства AP2, SUPERNUMERARY BRACT ( SNB ) и OsINDETERMINATE SPIKELET 1 ( OsIDS1 ), синергетически контролируют архитектуру соцветия и формирование меристемы цветков у риса. Plant J. 69, 445–461. DOI: 10.1111 / j.1365-313X.2011.04804.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ли, Д.Ю., Ли, Дж., Мун, С., Парк, С. Ю., и Ан, Г. (2007). Гетерохронный ген риса SUPERNUMERARY BRACT регулирует переход от меристемы колосков к меристеме цветков. Plant J. 49, 64–78. DOI: 10.1111 / j.1365-313X.2006.02941.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Lenhard, M., Bohnert, A., Jürgens, G., and Laux, T. (2001). Прекращение поддержания стволовых клеток в цветочных меристемах Arabidopsis посредством взаимодействий между WUSCHEL и AGAMOUS . Cell 105, 805–814. DOI: 10.1016 / S0092-8674 (01) 00390-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Li, H., Liang, W., Jia, R., Yin, C., Zong, J., Kong, H., et al. (2010). AGL6 -подобный ген OsMADS6 регулирует идентичность цветочных органов и меристем у риса. Cell Res. 20, 299–313. DOI: 10.1038 / cr.2009.143
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ли, Х., Сюэ, Д., Гао, З., Янь, М., Сюй, В., Син, З. и др. (2009). Предполагаемый ген липазы EXTRA GLUME1 регулирует судьбу пустой чешуи и развитие колосков у риса. Plant J. 57, 593–605. DOI: 10.1111 / j.1365-313X.2008.03710.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Луо, М., Платтен, Д., Чаудхури, А., Пикок, У. Дж., И Деннис, Э. С. (2009). Экспрессия, импринтинг и эволюция гомологов риса генов поликомбовой группы. Мол. Завод 2, 711–723.DOI: 10.1093 / mp / ssp036
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Майер, К. Ф., Шоф, Х., Хеккер, А., Ленхард, М., Юргенс, Г., и Лаукс, Т. (1998). Роль WUSCHEL в регуляции судьбы стволовых клеток в меристеме побегов Arabidopsis . Cell 95, 805–815. DOI: 10.1016 / S0092-8674 (00) 81703-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мун, С., Юнг, К. Х., Ли, Д. Э., Ли, Д. Ю., Ли, Дж., Ан, К. и др. (2006). Ген риса FON1 контролирует вегетативное и репродуктивное развитие, регулируя размер апикальной меристемы побега. Мол. Ячейки 21, 147–152.
PubMed Аннотация | Google Scholar
Мун, Ю. Х., Чен, Л., Пан, Р. Л., Чанг, Х. С., Чжу, Т., Маффео, Д. М. и др. (2003). Гены EMF поддерживают вегетативное развитие, подавляя программу цветков Arabidopsis . Растительная клетка 15, 681–693. DOI: 10.1105 / тп. 007831
CrossRef Полный текст | Google Scholar
Мозгова И., Кёлер К. и Хенниг Л. (2015). Держим ворота закрытыми: функции репрессивного комплекса polycomb PRC2 в разработке. Plant J. 83, 121–132. DOI: 10.1111 / tpj.12828
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мюллер, Дж., Харт, К. М., Фрэнсис, Н. Дж., Варгас, М. Л., Сенгупта, А., Уайлд, Б. и др. (2002). Гистон-метилтрансферазная активность репрессорного комплекса группы Drosophila Polycomb. Ячейка 111, 197–208. DOI: 10.1016 / S0092-8674 (02) 00976-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Накагава М., Симамото К. и Киодзука Дж. (2002). Избыточная экспрессия гомологов RCN1 и RCN2 риса TERMINAL FLOWER 1 / CENTRORADIALIS вызывает задержку фазового перехода и измененную морфологию метелки у риса. Plant J. 29, 743–750. DOI: 10.1046 / j.1365-313X.2002.01255.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Нардманн, Дж., и Верр, W. (2006). Ниша стволовых клеток побегов у покрытосеменных: паттерны экспрессии ортологов WUS у риса и кукурузы предполагают значительные модификации в ходе эволюции моно- и двудольных. Мол. Биол. Evol. 23, 2492–2504. DOI: 10.1093 / molbev / msl125
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Омори, С., Кимизу, М., Сугита, М., Мияо, А., Хирочика, Х., Учида, Э. и др. (2009). MOSAIC FLORAL ORGANS1 , AGL6 -подобный ген MADS box, регулирует идентичность цветочных органов и судьбу меристемы у риса. Растительная клетка 21, 3008–3025. DOI: 10.1105 / tpc.109.068742
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Прасад, К., Парамешваран, С., Виджайрагхаван, У. (2005). OsMADS1 , фактор MADS-бокса риса, контролирует дифференцировку определенных типов клеток в lemma и palea и является регулятором раннего действия внутренних органов цветков. Plant J. 43, 915–928. DOI: 10.1111 / j.1365-313X.2005.02504.x
PubMed Аннотация | CrossRef Полный текст
Рао, Н.Н., Прасад, К., Кумар, П. Р., Виджайрагаван, У. (2008). Определенная регулирующая роль RFL , гомолога риса LFY , в определении времени цветения и строения растений. Proc. Natl. Акад. Sci. США 105, 3646–3651. DOI: 10.1073 / pnas.0700
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ren, D., Li, Y., Zhao, F., Sang, X., Shi, J., Wang, N., et al. (2013). MULTI-FLORET SPIKELET1 , который кодирует белок AP2 / ERF, определяет судьбу меристемы колосков и идентичность стерильной леммы у риса. Plant Physiol. 162, 872–884. DOI: 10.1104 / PP.113.216044
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Sang, X., Li, Y., Luo, Z., Ren, D., Fang, L., Wang, N., et al. (2012). CHIMERIC FLORAL ORGANS1 , кодирующий белок MADS box, специфичный для однодольных растений, регулирует идентичность органов цветков риса. Plant Physiol. 160, 788–807. DOI: 10.1104 / стр.112.200980
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вс, Б., Looi, L. S., Guo, S., He, Z., Gan, E. S., Huang, J., et al. (2014). Механизм времени, зависящий от клеточного деления, запускается выселением Polycomb в стволовых клетках растений. Наука 343: 1248559. DOI: 10.1126 / science.1248559
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сунь, Б., Сюй, Ю., Нг, К. Х., Ито, Т. (2009). Механизм времени для поддержания и дифференцировки стволовых клеток в цветочной меристеме Arabidopsis . Genes Dev. 23, 1791–1804.DOI: 10.1101 / gad.1800409
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сунг, С., Шмитц, Р. Дж., И Амасино, Р. М. (2006). Пальцевый белок PHD, участвующий как в пути яровизации, так и в пути фотопериода у Arabidopsis . Genes Dev. 20, 3244–3248. DOI: 10.1101 / gad.14
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сузаки Т., Сато М., Асикари М., Миёши М., Нагато Ю. и Хирано Х. Ю.(2004). Ген FLORAL ORGAN NUMBER1 регулирует размер меристемы цветков у риса и кодирует киназу рецептора с высоким содержанием лейцина, ортологичную Arabidopsis CLAVATA1. Развитие 131, 5649–5657. DOI: 10.1242 / dev.01441
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wang, K., Tang, D., Hong, L., Xu, W., Huang, J., Li, M., et al. (2010). DEP и AFO регулируют репродуктивную функцию риса. PLOS Genet. 6: e1000818. DOI: 10.1371 / journal.pgen.1000818
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wei, J., Choi, H., Jin, P., Wu, Y., Yoon, J., Lee, Y.S, et al. (2016). Гомеобокса типа GL2 Roc4 в рисе способствует продолжительности цветения, предпочтительно в длинные дни, подавляя Ghd7 . Plant Sci. 252, 133–143. DOI: 10.1016 / j.plantsci.2016.7.012
CrossRef Полный текст | Google Scholar
Се, С., Чен, М., Пей, Р., Оуян, Ю., Яо, Дж. (2015). OsEMF2b действует как регулятор перехода цветения и идентичности цветковых органов, опосредуя отложение h4K27me3 на OsLFL1 и OsMADS4 в рисе. Завод Мол. Биол. Rep. 33, 121–132. DOI: 10.1007 / s11105-014-0733-1
CrossRef Полный текст | Google Scholar
Янг, Дж., Ли, С., Ханг, Р., Ким, С. Р., Ли, Ю. С., Цао, X., et al. (2013). OsVIL2 действует вместе с PRC2, вызывая цветение, подавляя OsLFL1 в рисе. Plant J. 73, 566–578. DOI: 10.1111 / tpj.12057
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Юн, Дж., Чо, Л. Х., Ким, С. Л., Чой, Х., Кох, Х. Дж. И Ан, Г. (2014). Ген гомеобокса типа BEL1 SH5 индуцирует раздробление семян за счет усиления развития зоны отслоения и ингибирования биосинтеза лигнина. Plant J. 79, 717–728. DOI: 10.1111 / tpj.12581
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ёсида, А., Сузаки, Т., Танака, В., Хирано, Х. Ю. (2009). Гомеотический ген длинная стерильная лемма ( G1 ) указывает идентичность стерильной леммы в рисовом колоске. Proc. Natl. Акад. Sci. США 106, 20103–20108. DOI: 10.1073 / pnas.0
6106PubMed Аннотация | CrossRef Полный текст | Google Scholar
Йошида, Н., Янаи, Ю., Чен, Л., Като, Ю., Хирацука, Дж., Мива, Т. и др. (2001). ЭМБРИОННЫЙ ЦВЕТОК2, новый гомолог белков группы поликомб, опосредует развитие и цветение побегов Arabidopsis . Растительная клетка 13, 2471–2481. DOI: 10.1105 / tpc.010227
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Юань, З., Гао, С., Сюэ, Д. В., Ло, Д., Ли, Л. Т., Дин, С. Ю. и др. (2009). RETARDED PALEA1 контролирует развитие палеа и цветочную зигоморфию риса. Plant Physiol. 149, 235–244. DOI: 10.1104 / стр.108.128231
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжан Д. и Юань З. (2014).Молекулярный контроль развития стеклянных соцветий. Annu. Rev. Plant Physiol. 65, 553–578. DOI: 10.1146 / annurev-arplant-050213-040104
PubMed Аннотация | CrossRef Полный текст
Zhang, J., Tang, W., Huang, Y., Niu, X., Zhao, Y., Han, Y., et al. (2015). Подавление гена LBD , OsIG1 , приводит к появлению необычных двойных семяпочек и аномалиям развития различных органов цветка и мегагаметофитов у риса. Дж.Exp. Бот. 66, 99–112. DOI: 10.1093 / jxb / eru396
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжао, С.К., Ху, Дж., Го, Л. Б., Цянь, К., и Сюэ, Х. У. (2010). Наклон листа риса2, белок, подобный VIN3, регулирует угол наклона листа посредством модуляции деления клеток воротничка. Cell Res. 20, 935–947. DOI: 10.1038 / cr.2010.109
PubMed Аннотация | CrossRef Полный текст | Google Scholar
.